FtsZ
FtsZ is a protein encoded by the ftsZ gene that assembles into a ring at the future site of bacterial cell division (also called the Z ring). FtsZ is a prokaryotic homologue of the eukaryotic protein tubulin. The initials FtsZ mean "Filamenting temperature-sensitive mutant Z." The hypothesis was that cell division mutants of E. coli would grow as filaments due to the inability of the daughter cells to separate from one another. FtsZ is found in almost all bacteria, many archaea, all chloroplasts and some mitochondria, where it is essential for cell division. FtsZ assembles the cytoskeletal scaffold of the Z ring that, along with additional proteins, constricts to divide the cell in two.
| Cell division protein FtsZ | |
|---|---|
| Identifiers | |
| Symbol | FtsZ |
| InterPro | IPR000158 |
| CATH | 1fsz |
| SCOP2 | 1fsz / SCOPe / SUPFAM |
| CDD | cd02201 |
| FtsZ, C-terminal sandwich | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||
| Symbol | FtsZ_C | ||||||||
| Pfam | PF12327 | ||||||||
| InterPro | IPR024757 | ||||||||
| |||||||||
| Cell division protein FtsZ | |||||||
|---|---|---|---|---|---|---|---|
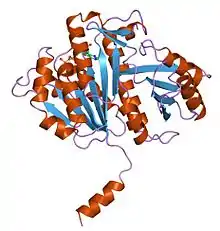 Molecular Structure of FtsZ (PDB 1fsz). | |||||||
| Identifiers | |||||||
| Organism | |||||||
| Symbol | ftsZ | ||||||
| UniProt | P0A9A6 | ||||||
| |||||||
History
In the 1960s scientists screened for temperature sensitive mutations that blocked cell division at 42 °C. The mutant cells divided normally at 30°, but failed to divide at 42°. Continued growth without division produced long filamentous cells (Filamenting temperature sensitive). Several such mutants were discovered and mapped to a locus originally named ftsA, which could be one or more genes. In 1980 Lutkenhaus and Donachie[1] showed that several of these mutations mapped to one gene, ftsA, but one well-characterized mutant, PAT84, originally discovered by Hirota et al,[2] mapped to a separate, adjacent gene. They named this cell division gene ftsZ. In 1991 Bi and Lutkenhaus used immunogold electron microscopy to show that FtsZ localized to the invaginating septum at midcell.[3] Subsequently, the Losick and Margolin groups used immuno-fluorescence microscopy[4] and GFP fusions[5] to show that FtsZ assembled Z rings early in the cell cycle, well before the septum began to constrict. Other division proteins then assemble onto the Z ring and constriction occurs in the last part of the cell cycle.
In 1992-3 three labs independently discovered that FtsZ was related to eukaryotic tubulin, which is the protein subunit that assembles into microtubules.[6][7][8] This was the first discovery that bacteria have homologs of eukaryotic cytoskeletal proteins. Later work showed that FtsZ was present in, and essential for, cell division in almost all bacteria and in many but not all archaea.
Mitochondria and chloroplasts are eukaryotic organelles that originated as bacterial endosymbionts, so there was much interest in whether they use FtsZ for division. Chloroplast FtsZ was first discovered by Osteryoung,[9] and it is now known that all chloroplasts use FtsZ for division. Mitochondrial FtsZ was discovered by Beech[10] in an alga; FtsZ is used for mitochondrial division in some eukaryotes, while others have replaced it with a dynamin-based machinery.
In 2014, scientists identified two FtsZ homologs in archaea, FtsZ1 and FtsZ2. [11]
Function

During cell division, FtsZ is the first protein to move to the division site, and is essential for recruiting other proteins that produce a new cell wall (septum) between the dividing cells. FtsZ's role in cell division is analogous to that of actin in eukaryotic cell division, but, unlike the actin-myosin ring in eukaryotes, FtsZ has no known motor protein associated with it. Cell wall synthesis may externally push the cell membrane, providing the force for cytokinesis. Supporting this, in E. coli the rate of division is affected by mutations in cell wall synthesis.[12] Alternatively, FtsZ may pull the membrane from the inside based on Osawa (2009) showing the protein's contractile force on liposomes with no other proteins present.[13]
Erickson (2009) proposed how the roles of tubulin-like proteins and actin-like proteins in cell division became reversed in an evolutionary mystery.[14] The use of the FtsZ ring in dividing chloroplasts and some mitochondria further establishes their prokaryotic ancestry.[15] L-form bacteria that lack a cell wall do not require FtsZ for division, which implies that bacteria may have retained components of an ancestral mode of cell division.[16]
Much is known about the dynamic polymerization activities of tubulin and microtubules, but little is known about these activities in FtsZ. While it is known that single-stranded tubulin protofilaments form into 13 stranded microtubules, the multistranded structure of the FtsZ-containing Z-ring is not known. It is only speculated that the structure consists of overlapping protofilaments. Nevertheless, recent work with purified FtsZ on supported lipid bilayers as well as imaging FtsZ in living bacterial cells revealed that FtsZ protofilaments have polarity and move in one direction by treadmilling[17] (see also below).
Recently, proteins similar to tubulin and FtsZ have been discovered in large plasmids found in Bacillus species. They are believed to function as components of segrosomes, which are multiprotein complexes that partition chromosomes/plasmids in bacteria. The plasmid homologs of tubulin/FtsZ seem to have conserved the ability to polymerize into filaments.
The contractile ring (the "Z ring")

FtsZ has the ability to bind to GTP and also exhibits a GTPase domain that allows it to hydrolyze GTP to GDP and a phosphate group. In vivo, FtsZ forms filaments with a repeating arrangement of subunits, all arranged head-to-tail.[18] These filaments form a ring around the longitudinal midpoint, or septum, of the cell. This ring is called the Z-ring.
The GTP hydrolyzing activity of the protein is not essential to the formation of filaments or cell division. Mutants defective in GTPase activity often still divide, but sometimes form twisted and disordered septa. It is unclear as to whether FtsZ actually provides the physical force that results in division or serves as a scaffold for other proteins to execute division.
There are two models for how FtsZ might generate a constriction force. One model is based on the observation that FtsZ protfilaments can be straight or curved. The transition from straight to curved is suggested to generate a bending force on the membrane.[19] Another model is based on sliding protofilaments. Computer models and in vivo measurements suggest that single FtsZ filaments cannot sustain a length more than 30 subunits long. In this model, FtsZ scission force comes from the relative lateral movement of subunits.[20] Lines of FtsZ would line up together parallel and pull on each other creating a "cord" of many strings that tightens itself.
In other models, FtsZ does not provide the contractile force but provides the cell a spatial scaffold for other proteins to execute the division of the cell. This is akin to the creating of a temporary structure by construction workers to access hard-to-reach places of a building. The temporary structure allows unfettered access and ensures that the workers can reach all places. If the temporary structure is not correctly built, the workers will not be able to reach certain places, and the building will be deficient.
The scaffold theory is supported by information that shows that the formation of the ring and localization to the membrane requires the concerted action of a number of accessory proteins. ZipA or the actin homologue FtsA permit initial FtsZ localization to the membrane.[21] Following localization to the membrane, division proteins of the Fts family are recruited for ring assembly.[22] Many of these proteins direct the synthesis of the new division septum at midcell (FtsI, FtsW), or regulate the activity of this synthesis (FtsQ, FtsL, FtsB, FtsN). The timing of Z-ring formation suggests the possibility of a spatial or temporal signal that permits the formation of FtsZ filaments.
Recent super-resolution imaging in several species supports a dynamic scaffold model, in which small clusters of FtsZ protofilaments or protofilament bundles move unidirectionally around the ring's circumference by treadmilling, anchored to the membrane by FtsA and other FtsZ-specific membrane tethers.[23][24] The speed of treadmilling depends on the rate of GTP hydrolysis within the FtsZ protofilaments, but in Escherichia coli, synthesis of the division septum remains the rate limiting step for cytokinesis.[25] The treadmilling action of FtsZ is required for proper synthesis of the division septum by septal peptidoglycan synthesis enzymes, suggesting that these enzymes can track the growing ends of the filaments.
Septal localization and intracellular signaling
The formation of the Z-ring closely coincides with cellular processes associated with replication. Z-ring formation coincides with the termination of genome replication in E. coli and 70% of chromosomal replication in B. subtilis.[26] The timing of Z-ring formation suggests the possibility of a spatial or temporal signal that permits the formation of FtsZ filaments. In Escherichia coli, at least two negative regulators of FtsZ assembly form a bipolar gradient, such that the concentration of active FtsZ required for FtsZ assembly is highest at mid-cell between the two segregating chromosomes, and lowest at the poles and over the chromosomes. This type of regulation seems to occur in other species such as Bacillus subtilis and Caulobacter crescentus. However, other species including Streptococcus pneumoniae and Myxococcus xanthus seem to use positive regulators that stimulate FtsZ assembly at mid-cell.[27]
Communicating distress
FtsZ polymerization is also linked to stressors like DNA damage. DNA damage induces a variety of proteins to be manufactured, one of them called SulA.[28] SulA prevents the polymerization and GTPase activity of FtsZ. SulA accomplishes this task by binding to self-recognizing FtsZ sites. By sequestering FtsZ, the cell can directly link DNA damage to inhibiting cell division.[29]
Preventing DNA damage
Like SulA, there are other mechanisms that prevent cell division that would result in disrupted genetic information sent to daughter cells. So far, two proteins have been identified in E. coli and B. subtilis that prevent division over the nucleoid region: Noc and SlmA. Noc gene knockouts result in cells that divide without respect to the nucleoid region, resulting in its asymmetrical partitioning between the daughter cells. The mechanism is not well understood, but thought to involve sequestration of FtsZ, preventing polymerization over the nucleoid region.[30] The mechanism used by SlmA to inhibit FtsZ polymerization over the nucleoid [31] is better understood, and uses two separate steps. One domain of SlmA binds to a FtsZ polymer, then a separate domain of SlmA severs the polymer.[32] A similar mechanism is thought to be used by MinC, another inhibitor of FtsZ polymerization involved in positioning of the FtsZ ring.[33]
Clinical significance
The number of multidrug-resistant bacterial strains is currently increasing; thus, the determination of drug targets for the development of novel antimicrobial drugs is urgently needed. The potential role of FtsZ in the blockage of cell division, together with its high degree of conservation across bacterial species, makes FtsZ a highly attractive target for developing novel antibiotics.[34] Researchers have been working on synthetic molecules and natural products as inhibitors of FtsZ.[35]
The spontaneous self-assembly of FtsZ can also be used in nanotechnology to fabricate metal nanowires.[36][37]
See also
- Fission (biology) – Biological process
- Divisome – A protein complex in bacteria responsible for cell division
- FtsK – Protein involved in bacterial cell division
References
- Lutkenhaus JF, Wolf-Watz H, Donachie WD (May 1980). "Organization of genes in the ftsA-envA region of the Escherichia coli genetic map and identification of a new fts locus (ftsZ)". Journal of Bacteriology. 142 (2): 615–620. doi:10.1128/JB.142.2.615-620.1980. OCLC 678550294. PMC 294035. PMID 6991482.
- Hirota Y, Ryter A, Jacob F (1968-01-01). "Thermosensitive mutants of E. coli affected in the processes of DNA synthesis and cellular division". Cold Spring Harbor Symposia on Quantitative Biology. 33: 677–693. doi:10.1101/sqb.1968.033.01.077. PMID 4892005.
- Bi EF, Lutkenhaus J (November 1991). "FtsZ ring structure associated with division in Escherichia coli". Nature. 354 (6349): 161–164. Bibcode:1991Natur.354..161B. doi:10.1038/354161a0. PMID 1944597. S2CID 4329947.
- Levin PA, Losick R (February 1996). "Transcription factor Spo0A switches the localization of the cell division protein FtsZ from a medial to a bipolar pattern in Bacillus subtilis". Genes & Development. 10 (4): 478–488. doi:10.1101/gad.10.4.478. PMID 8600030.
- Ma X, Ehrhardt DW, Margolin W (November 1996). "Colocalization of cell division proteins FtsZ and FtsA to cytoskeletal structures in living Escherichia coli cells by using green fluorescent protein". Proceedings of the National Academy of Sciences of the United States of America. 93 (23): 12998–13003. Bibcode:1996PNAS...9312998M. doi:10.1073/pnas.93.23.12998. PMC 24035. PMID 8917533.
- RayChaudhuri D, Park JT (September 1992). "Escherichia coli cell-division gene ftsZ encodes a novel GTP-binding protein". Nature. 359 (6392): 251–254. Bibcode:1992Natur.359..251R. doi:10.1038/359251a0. PMID 1528267. S2CID 4355143.
- de Boer P, Crossley R, Rothfield L (September 1992). "The essential bacterial cell-division protein FtsZ is a GTPase". Nature. 359 (6392): 254–256. Bibcode:1992Natur.359..254D. doi:10.1038/359254a0. PMID 1528268. S2CID 2748757.
- Mukherjee A, Dai K, Lutkenhaus J (February 1993). "Escherichia coli cell division protein FtsZ is a guanine nucleotide binding protein". Proceedings of the National Academy of Sciences of the United States of America. 90 (3): 1053–1057. Bibcode:1993PNAS...90.1053M. doi:10.1073/pnas.90.3.1053. PMC 45809. PMID 8430073.
- Osteryoung KW, Vierling E (August 1995). "Conserved cell and organelle division". Nature. 376 (6540): 473–474. Bibcode:1995Natur.376..473O. doi:10.1038/376473b0. PMID 7637778. S2CID 5399155.
- Beech PL, Nheu T, Schultz T, Herbert S, Lithgow T, Gilson PR, McFadden GI (February 2000). "Mitochondrial FtsZ in a chromophyte alga". Science. 287 (5456): 1276–1279. Bibcode:2000Sci...287.1276B. doi:10.1126/science.287.5456.1276. PMID 10678836. S2CID 26587576.
- Duggin, Iain G.; Aylett, Christopher H. S.; Walsh, James C.; Michie, Katharine A.; Wang, Qing; Turnbull, Lynne; Dawson, Emma M.; Harry, Elizabeth J.; Whitchurch, Cynthia B.; Amos, Linda A.; Löwe, Jan (2014-12-22). "CetZ tubulin-like proteins control archaeal cell shape". Nature. Springer Science and Business Media LLC. 519 (7543): 362–365. doi:10.1038/nature13983. ISSN 0028-0836. PMC 4369195.
- Coltharp C, Buss J, Plumer TM, Xiao J (February 2016). "Defining the rate-limiting processes of bacterial cytokinesis". Proceedings of the National Academy of Sciences of the United States of America. 113 (8): E1044–E1053. Bibcode:2016PNAS..113E1044C. doi:10.1073/pnas.1514296113. PMC 4776500. PMID 26831086.
- Osawa M, Anderson DE, Erickson HP (May 2008). "Reconstitution of contractile FtsZ rings in liposomes". Science. 320 (5877): 792–794. Bibcode:2008Sci...320..792O. doi:10.1126/science.1154520. PMC 2645864. PMID 18420899.
- Erickson HP (July 2007). "Evolution of the cytoskeleton". BioEssays. 29 (7): 668–677. doi:10.1002/bies.20601. PMC 2630885. PMID 17563102.
- Leger MM, Petrů M, Žárský V, Eme L, Vlček Č, Harding T, et al. (August 2015). "An ancestral bacterial division system is widespread in eukaryotic mitochondria". Proceedings of the National Academy of Sciences of the United States of America. 112 (33): 10239–10246. Bibcode:2015PNAS..11210239L. doi:10.1073/pnas.1421392112. PMC 4547283. PMID 25831547.
- Leaver M, Domínguez-Cuevas P, Coxhead JM, Daniel RA, Errington J (February 2009). "Life without a wall or division machine in Bacillus subtilis". Nature. 457 (7231): 849–853. Bibcode:2009Natur.457..849L. doi:10.1038/nature07742. PMID 19212404. S2CID 4413852.
- Loose M, Mitchison TJ (January 2014). "The bacterial cell division proteins FtsA and FtsZ self-organize into dynamic cytoskeletal patterns". Nature Cell Biology. 16 (1): 38–46. doi:10.1038/ncb2885. PMC 4019675. PMID 24316672.
- Desai A, Mitchison TJ (1997). "Microtubule polymerization dynamics". Annual Review of Cell and Developmental Biology. 13: 83–117. doi:10.1146/annurev.cellbio.13.1.83. PMID 9442869.
- Erickson HP, Anderson DE, Osawa M (December 2010). "FtsZ in bacterial cytokinesis: cytoskeleton and force generator all in one". Microbiology and Molecular Biology Reviews. 74 (4): 504–528. doi:10.1128/MMBR.00021-10. PMC 3008173. PMID 21119015.
- Lan G, Daniels BR, Dobrowsky TM, Wirtz D, Sun SX (January 2009). "Condensation of FtsZ filaments can drive bacterial cell division". Proceedings of the National Academy of Sciences of the United States of America. 106 (1): 121–126. Bibcode:2009PNAS..106..121L. doi:10.1073/pnas.0807963106. PMC 2629247. PMID 19116281.
- Pichoff S, Lutkenhaus J (March 2005). "Tethering the Z ring to the membrane through a conserved membrane targeting sequence in FtsA". Molecular Microbiology. 55 (6): 1722–1734. doi:10.1111/j.1365-2958.2005.04522.x. PMID 15752196.
- Buddelmeijer N, Beckwith J (December 2002). "Assembly of cell division proteins at the E. coli cell center". Current Opinion in Microbiology. 5 (6): 553–557. doi:10.1016/S1369-5274(02)00374-0. PMID 12457697.
- Yang X, Lyu Z, Miguel A, McQuillen R, Huang KC, Xiao J (February 2017). "GTPase activity-coupled treadmilling of the bacterial tubulin FtsZ organizes septal cell wall synthesis". Science. 355 (6326): 744–747. Bibcode:2017Sci...355..744Y. doi:10.1126/science.aak9995. PMC 5851775. PMID 28209899.
- Bisson-Filho AW, Hsu YP, Squyres GR, Kuru E, Wu F, Jukes C, et al. (February 2017). "Treadmilling by FtsZ filaments drives peptidoglycan synthesis and bacterial cell division". Science. 355 (6326): 739–743. Bibcode:2017Sci...355..739B. doi:10.1126/science.aak9973. PMC 5485650. PMID 28209898.
- Coltharp C, Buss J, Plumer TM, Xiao J (February 2016). "Defining the rate-limiting processes of bacterial cytokinesis". Proceedings of the National Academy of Sciences of the United States of America. 113 (8): E1044–E1053. Bibcode:2016PNAS..113E1044C. doi:10.1073/pnas.1514296113. PMC 4776500. PMID 26831086.
- Harry EJ (January 2001). "Coordinating DNA replication with cell division: lessons from outgrowing spores". Biochimie. 83 (1): 75–81. doi:10.1016/S0300-9084(00)01220-7. PMID 11254978.
- Rowlett VW, Margolin W (2015). "The Min system and other nucleoid-independent regulators of Z ring positioning". Frontiers in Microbiology. 6: 478. doi:10.3389/fmicb.2015.00478. PMC 4429545. PMID 26029202.
- He AS, Rohatgi PR, Hersh MN, Rosenberg SM (February 2006). "Roles of E. coli double-strand-break-repair proteins in stress-induced mutation". DNA Repair. 5 (2): 258–273. doi:10.1016/j.dnarep.2005.10.006. PMC 3685484. PMID 16310415.
- Mukherjee A, Lutkenhaus J (January 1998). "Dynamic assembly of FtsZ regulated by GTP hydrolysis". The EMBO Journal. 17 (2): 462–469. doi:10.1093/emboj/17.2.462. PMC 1170397. PMID 9430638.
- Wu LJ, Errington J (June 2004). "Coordination of cell division and chromosome segregation by a nucleoid occlusion protein in Bacillus subtilis". Cell. 117 (7): 915–925. doi:10.1016/j.cell.2004.06.002. PMID 15210112.
- Bernhardt TG, de Boer PA (May 2005). "SlmA, a nucleoid-associated, FtsZ binding protein required for blocking septal ring assembly over Chromosomes in E. coli". Molecular Cell. 18 (5): 555–564. doi:10.1016/j.molcel.2005.04.012. PMC 4428309. PMID 15916962.
- Du S, Lutkenhaus J (July 2014). "SlmA antagonism of FtsZ assembly employs a two-pronged mechanism like MinCD". PLOS Genetics. 10 (7): e1004460. doi:10.1371/journal.pgen.1004460. PMC 4117426. PMID 25078077.
- Shen B, Lutkenhaus J (March 2010). "Examination of the interaction between FtsZ and MinCN in E. coli suggests how MinC disrupts Z rings". Molecular Microbiology. 75 (5): 1285–1298. doi:10.1111/j.1365-2958.2010.07055.x. PMID 20132438.
- Casiraghi A, Suigo L, Valoti E, Straniero V (February 2020). "Targeting Bacterial Cell Division: A Binding Site-Centered Approach to the Most Promising Inhibitors of the Essential Protein FtsZ". Antibiotics. 9 (2): 69. doi:10.3390/antibiotics9020069. PMC 7167804. PMID 32046082.
- Ur Rahman M, Wang P, Wang N, Chen Y (August 2020). "A key bacterial cytoskeletal cell division protein FtsZ as a novel therapeutic antibacterial drug target". Bosnian Journal of Basic Medical Sciences. 20 (3): 310–318. doi:10.17305/bjbms.2020.4597. PMC 7416170. PMID 32020845.
- Ostrov N, Gazit E (April 2010). "Genetic engineering of biomolecular scaffolds for the fabrication of organic and metallic nanowires". Angewandte Chemie. 49 (17): 3018–3021. doi:10.1002/anie.200906831. PMID 20349481.
- Ostrov N, Fichman G, Adler-Abramovich L, Gazit E (January 2015). "FtsZ Cytoskeletal Filaments as a Template for Metallic Nanowire Fabrication". Journal of Nanoscience and Nanotechnology. 15 (1): 556–561. doi:10.1166/jnn.2015.9203. PMID 26328401.