HLA-DR7
HLA-DR7 (DR7) is a HLA-DR serotype that recognizes the DRB1*0701 to *0705 gene products.
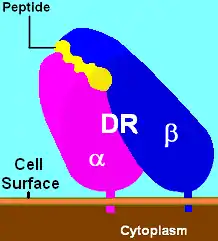 | |
human major histocompatibility complex, class II, DR7 | |
| Haplotypes groups | DQA*01:DRB1*0701 |
| Structure (See HLA-DR) | |
| Identifiers | alpha *01:01 |
| Symbol(s) | HLA-DRA |
| EBI-HLA | DRA*01:01 |
| Identifiers | beta 1 *07:01 |
| Symbol(s) | HLA-DRB1 |
| EBI-HLA | DRB1*07:01 |
| Shared data | |
| Locus | chr.6 6p21.31 |
Serology
| DRB1* | DR7 | Sample |
| allele | % | size (N) |
| 0701 | 99 | 7390 |
| 0704 | >30% | 1 |
The serological reaction of DR7 is excellent for *0701. The serology of *0703 to *0705 to *0709, and *0711 to *0714 serotypes is unknown. DRB1*0710N is a null allele. DRB1*0702 nomenclature has been deleted.
Disease associations
By serotype
DR7 is positively associated with psoriasis vulgaris.
By Allele group
DRB1*07 is linked to T. cruzi infection with cardiomyopathy (also called Chaga's cardiomyopathy).
By extended haplotype
DR7:DQA1*0201:DQB1*0202 is associated with Graves' disease.[2]
DR7-DQ2/DR5-DQ7 phenotype (transhaplotype encoded isoform DQα5β2) is the primary risk DQ isoform in celiac disease. DR7-DQ2/DR5-DQ7 (/DR11-DQ7 or /DR12-DQ7) is a clarifying identifier for the at risk transhaplotype.
DR53-DR7 may be associated with sclerosis/lupus associated anti-apolipoprotein antibodies.
Genetic linkage
DR7 Haplotypes | ||||
| Serotypes | DRA | DRB1 | DRB4 | |
|---|---|---|---|---|
| DR7-DR53 | *0101 | *0701 | *01 | |
| Serotypes | DQA1 | DQB1 | DRB1 | |
| DR7-DQ2 (2.2) | *0201 | *0202 | *0701 | |
| DR7-DQ9 (9.2, 3) | *0201 | *0303 | *0701 | |
| Serotypes | HLA-A | HLA-C | HLA-B | DRB1 |
| A29(19)-Cw16-B44(16)-DR7 | *2902 | *1601 | *4403 | *0701 |
| A30(19)-Cw6-B13-DR7 | *3001 | *0602 | *1302 | *0701 |
| A33(19)-Cw7-B44(16)-DR7 | *3301 | *0701 | *4403 | *0701 |
HLA-DR7 is genetically linked HLA-DR53, and is linked to DQ2 serotypes. There are a few interesting genetics with DR7. The A29-Cw16-B44-DR7-DQ2 haplotype is in strong linkage disequilibrium particularly in Northwestern Europe. The highest frequencies tend to be coastal countries along the Atlantic. The Cw16 allele is undoubtedly derived from Western Africa the diversity and frequency of Cw16 declines away from the along the Greenwich longitudinal line. The level of linkage disequilibration about Cw16 postulates a recent arrival from Africa, and indicates a substantial contribution as far north as Ireland. The A33-Cw7-B44-DR7 offers the same scenario. This haplotype is found in Korea but not Japan, in fact Japan is an antinode of DR7. The most frequent A-B-DR-DQ haplotypes in Korea are found at ~3/4 the Korean frequencies in Japan, except A33-Cw7-B44-DR7-DQ2, which indicates that DR7 likely spread into Korea after the Japan's Yayoi migration when most of the Korean contribution occurred. Similarities with Mongol genes suggest that Central and East- Central Asian are the likely source, confirmed by the fact the haplotype is found in Pakistan, China and India. There is the possibility that this haplotype is a recent (Holocene) arrival from Africa or the Middle East to Central Asia as this best explains the alleles within the haplotype. The A19 (A29, A30, A31, A32, A33, A74) serotypes are very much enriched in sub-saharan and Western Africa, including Iberia, and disequilibrated components of DR7 show a tendency to be linked to A19 serotypes.
References
- derived from IMGT/HLA
- Wongsurawat T, Nakkuntod J, Charoenwongse P, Snabboon T, Sridama V, Hirankarn N (2006). "The association between HLA class II haplotype with Graves' disease in Thai population". Tissue Antigens. 67 (1): 79–83. doi:10.1111/j.1399-0039.2005.00498.x. PMID 16451208.