Dinosaurios con plumas
Los dinosaurios con plumas se consideran formas de transición entre dinosaurios clásicos y aves. Era ya conocido que las aves antiguas como Archaeopteryx tenían muchas características de reptil, como dientes, y garras en sus dedos, y muchos años antes se había teorizado que los pájaros descendían de dinosaurios terópodos. A finales de los años 1990, los descubrimientos de dinosaurios con plumas en China proporcionaron la prueba concluyente de la conexión, aunque los detalles genealógicos todavía estén siendo dilucidados.
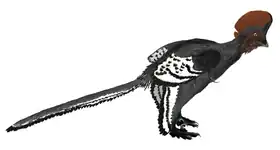
Se ha demostrado de muchas formas la relación de descendencia entre aves y dinosaurios, ya que su similitud morfológica en cuanto al cráneo, la cadera, las patas y las extremidades superiores es muy notable. Las aves son claramente monofiléticas y sus primeros representantes se encuentran en el Jurásico (Protoavis, un taxón controvertido de finales del Triásico, es considerado no aviar por la mayoría de los paleontólogos).
En 2017, Baron, Norman y Barrett propusieron que las plumas o las estructuras similares a estas pueden haberse originado en el ancestro común de los Ornithoscelida, un grupo de dinosaurios que incluiría tanto a los terópodos como a los ornitisquios, los dos únicos clados de dinosaurios en los cuales se ha observado la presencia de plumas.[1] Es incluso posible que las plumas se hubieran desarrollado en algún grupo anterior, a la luz del descubrimiento de las picnofibras de los pterosaurios. Los crocodilianos también poseen una beta-queratina muy similar a las de las aves, lo cual sugiere que su integumento evolucionó a partir de genes ancestrales comunes.[2]
Géneros de dinosaurios con plumas
Desde 1996 hasta hoy se han descrito más de 30 géneros de dinosaurios no aviares con plumas o protoplumas preservadas en sus fósiles:
- Avimimus* (inferido en 1987)[3][4]
- Sinosauropteryx (1996)[5]
- Protarchaeopteryx (1997)[6]
- Caudipteryx (1998)[7]
- Rahonavis* (1998)[8]
- Shuvuuia* (1999)[9]
- Sinornithosaurus (1999)[10]
- Beipiaosaurus (1999)[11]
- Microraptor (2000)[12]
- Nomingia* (2000)[13]
- Cryptovolans (2002)[14]
- Scansoriopteryx (2002)[15]
- Epidendrosaurus (2002)[16]
- Psittacosaurus sp.? (2002)[17]
- Yixianosaurus (2003)[18]
- Dilong (2004)[19]
- Pedopenna (2005)[20]
- Jinfengopteryx (2005)[21][22]
- Juravenator (2006)[23][24]
- Sinocalliopteryx (2007)[25]
- Velociraptor* (inferido en 2007: protuberancias de cañones de plumas)[26]
- Epidexipteryx (2008)[27]
- Similicaudipteryx (inferido en 2008: pigóstilo; confirmado en 2010)[28][29]
- Anchiornis (2009)[30]
- ¿Tianyulong? (2009)[31]
- Concavenator corcovatus? (inferido en 2010)[32]
- Yutyrannus (2012)[33]
- Sciurumimus (2012)[34]
- Ornithomimus (2012)[35]
- Ningyuansaurus (2012)[36]
- Eosinopteryx (2013)[37]
- Citipati (inferido en 2013: pigóstilo)[38]
- Conchoraptor (infererido en 2013: pigóstilo)[38]
- Jianchangosaurus (2013)[39]
- Aurornis (2013; posible avial)[40]
- Changyuraptor (2014)[41]
- ¿Kulindadromeus? (2014)[42]
- Citipati (inferido 2014: pigóstilo)[43]
- Conchoraptor (inferido 2014: pigóstilo)[38]
- Deinocheirus (inferido en 2014: pigóstilo)[44]
- Yi qi (2015)[45]
- Zhenyuanlong (2015)[46]
- Dakotaraptor (inferido en 2015: protuberancias de cañones de plumas)[47]
- Apatoraptor (inferido en 2016: protuberancias de cañones de plumas)[48]
- Jianianhualong tengi (2017)[49][50]
- Serikornis sungei (2017)[51]
- Caihong (2018)[52]
- Nótese que las estructuras filamentosas halladas en algunos dinosaurios ornitisquios (Psittacosaurus, Tianyulong, Kulindadromeus) y los pterosaurios pueden o no ser homólogas con las plumas y protoplumas de los terópodos.[31][53]
Plumas

Archaeopteryx, el primer dinosaurio con plumas o protoave encontrado fue descubierto en 1861. El espécimen original fue encontrado en las calizas de Solnhofen en Alemania meridional, una destacable y poco común formación geológica conocida por sus extraordinariamente bien conservados fósiles. Archaeopteryx es una forma transicional con características claramente atribuibles a reptiles y a aves por igual. Hallado dos años después de la publicación de El origen de las especies de Charles Darwin, su descubrimiento y probable rol de "eslabón perdido" catapultó el debate entre evolucionistas y creacionistas. Esta ave primitiva es tan similar a un dinosaurio típico que —cuando las plumas no se preservaban en las improntas fósiles— sus ejemplares fueron confundidos con individuos del género Compsognathus.
Desde los años 1990 se ha encontrado un número creciente de dinosaurios con plumas, lo cual proveyó evidencia adicional de la cercana relación entre dinosaurios y aves. Los fósiles involucrados presentan rasgos de aves, incluyendo plumas que no son exactamente como las de las aves actuales, sino que reúnen algunas similitudes intermedias entre el pelo y el plumón, útiles para cubrirse del frío. Este tipo de pluma, más conocida como "protopluma", se considera precursora del tipo que las aves modernas poseen. Entre los dinosaurios con plumas descubiertos hasta ahora están Beipiaosaurus, Caudipteryx, Dilong, Microraptor, Protarchaeopteryx, Shuvuuia, Sinornithosaurus, Sinosauropteryx y Jinfengopteryx. Yutyrannus, descubierto en 2012, es el dinosaurio más grande que se conoce hasta ahora cubierto de protoplumas.
En los dromeosáuridos en particular parece generalizado la cubierta de plumas, y poseían plumas de notable complejidad. Y al menos un integrante de esta familia, Microraptor, podría haber sido capaz de planear.
La mayoría de estos especímenes con plumas han sido exhumados en la provincia de Liaoning en el noreste de China, que fue parte de un continente aislado durante el Cretáceo. A pesar de que rastros de plumas han sido encontrados solo en la formación Yixian y otros pocos sitios, es probable que dinosaurios en otras partes del mundo también hayan estado cubiertos por plumas.
La falta de evidencia fósil a escala planetaria de plumaje puede deberse al hecho de que las estructuras delicadas como piel, pelo y plumas no son preservadas por las usuales condiciones de fosilización, necesitando condiciones especiales del sustrato para ser conservado, lo que dificulta su presencia en el registro fósil.
Esqueleto

Los dinosaurios y las aves comparten muchas características: se han identificado más de cien aspectos anatómicos aviares en los esqueletos de los terópodos maniraptores , y son aceptados ampliamente como sus predecesores y parientes más cercanos. Algunos de estos rasgos en común se presentan en el cuello, pubis, muñeca, extremidades superiores, omóplato, cerco y huesos pectorales. En especial la fúrcula, hueso resultante de fusionar ambas clavículas, único en terópodos y aves .[54]
Todo esto afirma la idea de que los dinosaurios fueron efectivamente los antepasados de las aves, a cuya forma convergieron en un largo proceso de adaptaciones anatómicas y fisiológicas. La relación entre aves y dinosaurios y el proceso de evolución de su vuelo es algo complejo, y todavía se discute si se produjo por medio de dinosaurios arborícolas planeadores o por rápidos corredores que emprenderían el vuelo en los cambios de rasante del terreno.
En un principio la creencia generalizada entre los científicos es que los dinosaurios han dado origen a las aves en una progresión lineal directa; pero algunos científicos, como Gregory S. Paul, han presentado la hipótesis de que algún grupo, como los dromaeosáuridos, pudieron haber evolucionado en el sentido inverso, a partir de las aves, perdiendo la habilidad de volar pero manteniendo sus plumas de manera similar a las actuales avestruces.
Similitudes reproductivas
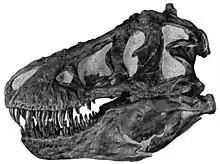
Un reciente descubrimiento efectuado en un esqueleto de Tyrannosaurus rex suministró pruebas adicionales de que los dinosaurios y las aves evolucionaron a partir de un ancestro común relativamente próximo y, por primera vez, permitió a los paleontólogos establecer el sexo de un dinosaurio. Cuando depositan sus huevos, las aves hembras desarrollan un tipo especial de tejido óseo en sus extremidades, llamado hueso medular, rico en calcio, que forma una capa dentro del hueso duro exterior, y que sirve para formar la cáscara del huevo. La presencia de esta clase de tejido óseo en el tuétano de porciones de los restos de las patas traseras del Tyrannosaurus sugiere que este animal usó estrategias reproductivas similares.
Pulmones
Los grandes dinosaurios carnívoros tuvieron un sistema de sacos de aire similar al que se encuentra en las aves modernas, de acuerdo a una investigación dirigida por Patrick O'Connor, de la Universidad de Ohio. Los pulmones de los terópodos probablemente empujaban aire hacia sacos vacíos en sus esqueletos, como en el caso de las aves. "Lo que una vez fue formalmente considerado exclusivo de las aves estuvo presente de alguna forma en sus ancestros", declaró O'Connor. El estudio fue parcialmente financiado por la Fundación Nacional para la Ciencia estadounidense.[55]
El corazón y la postura durante el sueño
Análisis de cavidades pectorales de dinosaurios hechas con tomografía computarizada en 2000 revelaron aparentes remanentes de complejos corazones de cuatro cavidades, parecidos a los que hoy tienen los mamíferos y las aves. Un fósil de Troodon recientemente descubierto demuestra que los dinosaurios durmieron como ciertas aves actuales, con sus cabezas escondidas bajo los brazos, postura que ayuda a conservar el calor craneal.[56] Este comportamiento pudo haber ayudado a mantener la cabeza caliente.
Gastrolitos
Otra similitud entre los dinosaurios y las aves es la ingestión de piedras con fines digestivos, que son tragadas por estos animales y ayudan a triturar las fibras resistentes de la comida cuando entran al estómago. Encontradas en asociación con fósiles, estas rocas son denominadas gastrolitos. Debido a que piedras identificables como pertenecientes a cierta formación geológica pueden haber sido engullidas en un lugar y trasladadas dentro del animal en sus desplazamientos, los paleontólogos a veces las usan para establecer posibles rutas de migración.
Taxonomía
Coelurosauria ├─Tugulusaurus (recubrimiento desconocido) │ └───Tyrannosauroidea │ ├──?Dilong paradoxus (impresiones de protoplumas) │ ├───Yutyrannus (impresiones de protoplumas) │ └───Tyrannosaurus (impresiones de escamas, probable pérdida secundaria de las protoplumas) │ ├─Compsognathidae │ ├─?Sinosauropteryx (impresiones de protoplumas) │ ├─Juravenator (impresiones de escamas, posible pérdida parcial o completa de las protoplumas) │ └─Compsognathus (recubrimiento desconocido) │ └─Maniraptoriformes ├─Ornithomimosauria │ └─Ornithomimus (impresiones de plumas complejas) ├─Alvarezsauridae │ └───Shuvuuia (plumas con cañón central y barbas; impresiones conservadas en 3 dimensiones) │ └───Maniraptora ├─Yixianosaurus (impresiones de plumas complejas) │ ├─Oviraptorosauria │ ├─Protarchaeopteryx (impresiones de plumas complejas) │ ├─Caudipteryx (impresiones de plumas complejas) │ └─Oviraptoridae (indicios en el esqueleto de presencia de plumas) │ │ │ ├─Therizinosauria │ └─Beipiaosaurus (impresiones de protoplumas) │ └───Paraves ├─Pedopenna (impresiones de plumas complejas) │ ├─Deinonychosauria │ ├─Troodontidae │ │ └─Jinfengopteryx (impresiones de plumas complejas) │ │ │ └─Dromaeosauridae │ ├─Rahonavis (esqueleto con indicios de plumas y adaptaciones anatómicas al vuelo) │ ├─Cryptovolans (impresiones de plumas complejas) │ ├─Microraptor (impresiones de plumas complejas. Alas con plumas asimétricas) │ ├─Sinornithosaurus (impresiones de protoplumas y plumas) │ └───Velociraptor (pruebas osteológicas de la presencia de plumas) │ ├─Epidendrosaurus (impresiones de plumas complejas) │ └───Aves (plumas complejas. Alas con plumas asimétricas)
Referencias
- Baron, M.G.; Norman, D.B.; Barrett, P.M. (2017). «A new hypothesis of dinosaur relationships and early dinosaur evolution». Nature 543: 501-506. doi:10.1038/nature21700.
- Greenwold, M.; Sawyer, R. (2013), «Molecular evolution and expression of archosaurian β-keratins: Diversification and expansion of archosaurian β-keratins and the origin of feather β-keratins», Journal of Experimental Zoology 320 (6): 393-405, PMID 23744807, doi:10.1002/jez.b.22514, 22514.
- Kurzanov, S.M. (1987). "Avimimidae and the problem of the origin of birds." Transactions of the Joint Soviet-Mongolian Paleontological Expedition, 31: 5-92. [in Russian]
- Chiappe, L.M. and Witmer, L.M. (2002). Mesozoic Birds: Above the Heads of Dinosaurs. Berkeley: University of California Press, 536 pp. ISBN 0-520-20094-2
- Ji, Q., and Ji, S. (1996). "On discovery of the earliest bird fossil in China and the origin of birds." Chinese Geology 10 (233): 30-33.
- Ji, Q., and Ji, S. (1997). "A Chinese archaeopterygian, Protarchaeopteryx gen. nov." Geological Science and Technology (Di Zhi Ke Ji), 238: 38-41. Translated By Will Downs Bilby Research Center Northern Arizona University January, 2001
- Qiang, J., .Currie, P.J., Norell., M.A. & Shu-An, J., 1998. Two feathered dinosaursfrom northeastern China. Nature 393 753-761.
- Forster, Catherine A.; Sampson, Scott D.; Chiappe, Luis M. & Krause, David W. (1998a). «The Theropod Ancestry of Birds: New Evidence from the Late Cretaceous of Madagascar». Science (5358): pp. 1915-1919.. doi 10.1126/science.279.5358.1915 (HTML abstract)
- Schweitzer, Mary Higby, Watt, J.A., Avci, R., Knapp, L., Chiappe, L, Norell, Mark A., Marshall, M. (1999). "Beta-Keratin Specific Immunological reactivity in Feather-Like Structures of the Cretaceous Alvarezsaurid, Shuvuuia deserti Journal of Experimental Biology (Mol Dev Evol) 255:146-157
- Xu, X., Wang, X., Wu, X., 1999. A dromaeosaurid dinosaur with a filamentous integument from the Yixian Formation of China. Nature 401:6750 262-266 doi 10.1038/45769
- Xu. X., Zhao, X., Clark, J.M., 1999. A new therizinosaur from the Lower Jurassic lower Lufeng Formation of Yunnan, China. Journal of Vertebrate Paleontology 21:3 477–483 doi 10.1671/0272-4634
- Xu, X., Zhou, Z., and Wang, X. (2000). "The smallest known non-avian theropod dinosaur." Nature, 408 (December): 705-708.
- Barsbold, R., Osmólska, H., Watabe, M., Currie, P.J., and Tsogtbaatar, K. (2000). "New Oviraptorosaur (Dinosauria, Theropoda) From Mongolia: The First Dinosaur With A Pygostyle". Acta Palaeontologica Polonica, 45(2): 97-106.
- Czerkas, S.A., Zhang, D., Li, J., and Li, Y. (2002). «Flying Dromaeosaurs». En Czerkas, S.J., ed. Feathered Dinosaurs and the Origin of Flight: The Dinosaur Museum Journal 1. Blanding: The Dinosaur Museum. pp. 16-26.
- Czerkas, S.A., and Yuan, C. (2002). "An arboreal maniraptoran from northeast China." Pp. 63-95 in Czerkas, S.J. (Ed.), Feathered Dinosaurs and the Origin of Flight. The Dinosaur Museum Journal 1. The Dinosaur Museum, Blanding, U.S.A. PDF abridged version
- Zhang, F., Zhou, Z., Xu, X. & Wang, X. (2002). "A juvenile coelurosaurian theropod from China indicates arboreal habits." Naturwissenschaften, 89(9): 394-398. doi=10.1007 /s00114-002-0353-8.
- Mayr, Gerald; Peters, Stefan; Plodowski, Gerhard; Vogel, Olaf (2002). «Bristle-like integumentary structures at the tail of the horned dinosaur Psittacosaurus». Naturwissenschaften 89 (8): 361-365. Bibcode:2002NW.....89..361M. PMID 12435037. doi:10.1007/s00114-002-0339-6.
- Xu, X. and Wang, X.-L. (2003). "A new maniraptoran from the Early Cretaceous Yixian Formation of western Liaoning." Vertebrata PalAsiatica, 41(3): 195–202.
- Xu, X., Norell, M. A., Kuang, X., Wang, X., Zhao, Q., Jia, C. (2004). «Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids». Nature 431: 680-684. doi:10.1038/nature02855. Archivado desde el original el 27 de marzo de 2009. Consultado el 4 de enero de 2009.
- Xu, X., and Zhang, F. (2005). "A new maniraptoran dinosaur from China with long feathers on the metatarsus." Naturwissenschaften, 92(4): 173 - 177.
- Ji, Q., Ji, S., Lu, J., You, H., Chen, W., Liu, Y., and Liu, Y. (2005). "First avialan bird from China (Jinfengopteryx elegans gen. et sp. nov.)." Geological Bulletin of China, 24(3): 197-205.
- Turner, Alan H.; Pol, Diego; Clarke, Julia A.; Erickson, Gregory M.; Norell, Mark (2007). «A basal dromaeosaurid and size evolution preceding avian flight» (pdf). Science 317: 1378-1381. PMID 17823350. doi:10.1126/science.1144066.
- Goehlich U.B., Tischlinger H., Chiappe L.M. (2006). «Juravenator starki (Reptilia, Theropoda) ein nuer Raubdinosaurier aus dem Oberjura der Suedlichen Frankenalb (Sueddeutschland): Skelettanatomie und Wiechteilbefunde». Archaeopteryx 24: 1-26.
- Chiappe, Luis M.; Göhlich, Ursula B. (2010). «Anatomy of Juravenator starki (Theropoda: Coelurosauria) from the Late Jurassic of Germany». Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen 258 (3): 257-296. doi:10.1127/0077-7749/2010/0125.
- Ji, S., Ji, Q., Lu J., and Yuan, C. (2007). "A new giant compsognathid dinosaur with long filamentous integuments from Lower Cretaceous of Northeastern China." Acta Geologica Sinica, 81(1): 8-15.
- Turner, A.H.; Makovicky, P.J.; Norell, M.A. (2007). «Feather quill knobs in the dinosaur Velociraptor» (pdf). Science 317 (5845): 1721. PMID 17885130. doi:10.1126/science.1145076.
- Zhang, F., Zhou, Z., Xu, X., Wang, X., & Sullivan, C. (2008). "A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers." Available from Nature Precedings, doi:10.1038/npre.2008.2326.1 .
- He T., Wang X.-L., Zhou Z.-H. (2008). «A new genus and species of caudipterid dinosaur from the Lower Cretaceous Jiufotang Formation of western Liaoning, China». Vertebrata PalAsiatica 46 (3): 178-189.
- Xu, Xing; Zheng, Xiaoting; You, Hailu (2010). «Exceptional dinosaur fossils show ontogenetic development of early feathers». Nature 464 (7293): 1338-1341. Bibcode:2010Natur.464.1338X. PMID 20428169. doi:10.1038/nature08965.
- Xu, X., Zhao, Q., Norell, M., Sullivan, C., Hone, D., Erickson, G., Wang, X., Han, F. and Guo, Y. (in press). "A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin." Chinese Science Bulletin, 6 pages, accepted November 15, 2008.
- Zheng, Xiao-Ting; You, Hai-Lu; Xu, Xing; Dong, Zhi-Ming (2009). «An Early Cretaceous heterodontosaurid dinosaur with filamentous integumentary structures». Nature 458 (7236): 333-336. Bibcode:2009Natur.458..333Z. PMID 19295609. doi:10.1038/nature07856.
- Ortega, Francisco; Escaso, Fernando; Sanz, José L. (2010). «A bizarre, humped Carcharodontosauria (Theropoda) from the Lower Cretaceous of Spain». Nature 467 (7312): 203-206. Bibcode:2010Natur.467..203O. PMID 20829793. doi:10.1038/nature09181.
- Xu, X., Wang, K., Zhang, K., Ma, Q., Xing, L., Sullivan, C., Hu, D., Cheng, S., and Wang, S. (2012). "A gigantic feathered dinosaur from the Lower Cretaceous of China. Archivado el 17 de abril de 2012 en Wayback Machine." Nature, 484: 92-95. doi 10.1038/nature10906
- Rauhut, O. W. M.; Foth, C.; Tischlinger, H.; Norell, M. A. (2012). «Exceptionally preserved juvenile megalosauroid theropod dinosaur with filamentous integument from the Late Jurassic of Germany». Proceedings of the National Academy of Sciences 109 (29): 11746. doi:10.1073/pnas.1203238109.<
- Darla K. Zelenitsky, François Therrien, Gregory M. Erickson, Christopher L. DeBuhr, Yoshitsugu Kobayashi, David A. Eberth, and Frank Hadfield. Feathered Non-Avian Dinosaurs from North America Provide Insight into Wing Origins. Science 26 October 2012: 338 (6106), 510-514. [DOI:10.1126/science.1225376]
- Ji Qiang, Lü Jun-Chang, Wei Xue-Fang, Wang Xu-Ri (2012). «A new oviraptorosaur from the Yixian Formation of Jianchang, Western Liaoning Province, China». Regional Geology of China (12): 2102-2107.
- Pascal Godefroit, Helena Demuynck, Gareth Dyke, Dongyu Hu, François Escuillié and Philippe Claeys (2013). «Reduced plumage and flight ability of a new Jurassic paravian theropod from China». Nature Communications 4: Article number 1394. doi:10.1038/ncomms2389.
- W. Scott Persons, IV, Philip J. Currie, and Mark A. Norell (in press). «Oviraptorosaur tail forms and functions». Acta Palaeontologica Polonica. doi:10.4202/app.2012.0093.
- Hanyong Pu, Yoshitsugu Kobayashi, Junchang Lü, Li Xu, Yanhua Wu, Huali Chang, Jiming Zhang, Songhai Jia (2013). «An unusual basal therizinosaur dinosaur with an ornithischian dental arrangement from northeastern China». PLOS ONE 8 (5): Article number e63423. Bibcode:2013PLoSO...863423P. doi:10.1371/journal.pone.0063423.
- Pascal Godefroit, Andrea Cau, Hu Dong-Yu, François Escuillié, Wu Wenhao, and Gareth Dyke (2013). «A Jurassic avialan dinosaur from China resolves the early phylogenetic history of birds». Nature 498: 359-362. doi:10.1038/nature12168.
- Gang Han, Luis M. Chiappe,Shu-An Ji, Michael Habib,Alan H. Turner, Anusuya Chinsamy, Xueling Liu, Lizhuo Han (2014). «A new raptorial dinosaur with exceptionally long feathering provides insights into dromaeosaurid flight performance». Nature Communications 5: Article number 4382. doi:10.1038/ncomms5382.
- Pascal Godefroit, Sofia M. Sinitsa, Danielle Dhouailly, Yuri L. Bolotsky, Alexander V. Sizov, Maria E. McNamara, Michael J. Benton, Paul Spagna (2014). «A Jurassic ornithischian dinosaur from Siberia with both feathers and scales». Science 25 (345): 451-455. doi:10.1126/science.1253351.
- W. Scott Persons, IV, Philip J. Currie, and Mark A. Norell (2014). «Oviraptorosaur tail forms and functions». Acta Palaeontologica Polonica. doi:10.4202/app.2012.0093.
- Yuong-Nam Lee, Rinchen Barsbold, Philip J. Currie, Yoshitsugu Kobayashi, Hang-Jae Lee, Pascal Godefroit, François Escuillié, and Tsogtbaatar Chinzorig (2014). «Resolving the long-standing enigmas of a giant ornithomimosaur Deinocheirus mirificus». Nature. doi:10.1038/nature13874.
- Xu, X.; Zheng, X.; Sullivan, C.; Wang, X.; Xing, L.; Wang, Y.; Zhang, X.; o’Connor, J. K.; Zhang, F.; Pan, Y. (2015). "A bizarre Jurassic maniraptoran theropod with preserved evidence of membranous wings". Nature.
- Lü, Junchang, Brusatte, Stephen L. 2015. A large, short-armed, winged dromaeosaurid (Dinosauria: Theropoda) from the Early Cretaceous of China and its implications for feather evolution. Sci. Rep.
- Robert A. DePalma, David A. Burnham, Larry D. Martin, Peter L. Larson and Robert T. Bakker (2015). "The first giant raptor (Theropoda: Dromaeosauridae) from the Hell Creek Formation". Paleontological Contributions 14: 1–16
- Gregory F. Funston; Philip J. Currie (2016). «A new caenagnathid (Dinosauria: Oviraptorosauria) from the Horseshoe Canyon Formation of Alberta, Canada, and a reevaluation of the relationships of Caenagnathidae». Journal of Vertebrate Paleontology. Online edition (4): e1160910. doi:10.1080/02724634.2016.1160910.
- Xing Xu; Philip Currie; Michael Pittman; Lida Xing; Qingjin Meng; Junchang Lü; Dongyu Hu; Congyu Yu (2017). «Mosaic evolution in an asymmetrically feathered troodontid dinosaur with transitional features». Nature Communications 8: Article number 14972. doi:10.1038/ncomms14972.
- «New Birdlike Dinosaur Had Modern Feathers». 2 de mayo de 2017. Consultado el 3 de mayo de 2017.
- Ulysse Lefèvre; Andrea Cau; Aude Cincotta; Dongyu Hu; Anusuya Chinsamy; François Escuillié; Pascal Godefroit (2017). «A new Jurassic theropod from China documents a transitional step in the macrostructure of feathers». The Science of Nature 104: Article number 74. doi:10.1007/s00114-017-1496-y.
- Dongyu Hu; Julia A. Clarke; Chad M. Eliason; Rui Qiu; Quanguo Li; Matthew D. Shawkey; Cuilin Zhao; Liliana D’Alba; Jinkai Jiang; Xing Xu (2018). «A bony-crested Jurassic dinosaur with evidence of iridescent plumage highlights complexity in early paravian evolution». Nature Communications 9: Article number 217. doi:10.1038/s41467-017-02515-y.
- Czerkas, S.A., and Ji, Q. (2002). "A new rhamphorhynchoid with a headcrest and complex integumentary structures." Pp. 15-41 in: Czerkas, S.J. (Ed.). Feathered Dinosaurs and the Origin of Flight. Blanding, Utah: The Dinosaur Museum. ISBN 1-932075-01-1.
- Mayr, G., Pohl, B. y Peters, D.S. A Well-Preserved Archaeopteryx Specimen with Theropod Features. Science 310:1483-1486. (2005).Véase comentario
- O'Connor, P.M. y Claessens, L.P.A.M. Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs. Nature 436:253. (2005).
- Xu, X. y Norell, M.A. A new troodontid dinosaur from China with avian-like sleeping posture. Nature 431:838-841. (2004).Véase comentario.
| Control de autoridades |
|
|---|
 Datos: Q828371
Datos: Q828371