Cretácico
El Cretácico, o Cretáceo, es una división de la escala temporal geológica que pertenece a la Era Mesozoica; dentro de esta, el Cretácico ocupa el tercer y último lugar siguiendo al Jurásico. Comenzó hace 145 millones de años y terminó hace 66 millones de años.[2] Con una duración de unos 79 millones de años, es el período Fanerozoico más extenso, y es, incluso, más largo que toda la Era Cenozoica. Su nombre proviene del latín creta, que significa "tiza",[3] y fue definido como un período independiente por el geólogo belga Jean d'Omalius d'Halloy en 1822, basándose en estratos de la cuenca parisina (Francia).[4]
| Era[1] | Período | Millones años |
|---|---|---|
| Mesozoica | Cretácico | 145,0 |
| Jurásico | 201,3 ±0,2 | |
| Triásico | 251,0 ±0,4 |

El Cretácico está dividido en dos grandes subunidades: Cretácico inferior y Cretácico superior.
La vida en mares y tierra aparecía como una mezcla de formas modernas y arcaicas, sobre todo de ammonites. Como ocurre con la mayoría de las eras geológicas, el inicio del período es incierto por unos pocos millones de años. Sin embargo, la datación del final del período es relativamente precisa, pues esta se hace coincidir con la de una capa geológica con fuerte presencia de iridio, que parece coincidir con la caída de un meteorito en lo que ahora corresponde con la península de Yucatán y el golfo de México. Este impacto pudo provocar la extinción masiva que ocurrió al final de este período, conocida como el evento K/Pg. Este acontecimiento marca el fin de la Era Mesozoica.
A mediados del Cretácico, se dio la formación de más del 50 % de las reservas mundiales de petróleo que se conocen en nuestros días, de las cuales se destacan las concentraciones localizadas en los alrededores del golfo Pérsico y en la región entre el golfo de México y la costa de Venezuela.
Divisiones
A continuación se indican las edades o pisos, ordenados «estratigráficamente», con las más recientes arriba y las más antiguas abajo:[5]
| Era Eratema |
Periodo Sistema |
Época Serie |
Edad Piso |
Inicio, en millones de años[6] |
|---|---|---|---|---|
| Mesozoico | Cretácico | Superior / Tardío | Maastrichtiense | |
| Campaniense | ||||
| Santoniense | ||||
| Coniaciense | ||||
| Turoniense | ||||
| Cenomaniense | ||||
| Inferior / Temprano | Albiense | |||
| Aptiense | ~121,4 | |||
| Barremiense | ||||
| Hauteriviense | ||||
| Valanginiense | ~139,8 | |||
| Berriasiense | ~145,0 | |||
| Jurásico | 201,4±0,2 | |||
| Triásico | 251,9±0,024 | |||
Estratigrafía
Península ibérica
En la península ibérica, el Cretácico es el período mesozoico que posee mayor extensión. En algunas zonas, la franja sedimentaria puede alcanzar hasta 2 kilómetros de grosor.[7] Los sedimentos que pertenecen al Cretácico inferior suelen ser detríticos (pudingas y areniscas); hacia la mitad del periodo aparecen margas, y finalmente se desarrollan calizas marinas, con alguna intercalación lacustre. En la Comunidad Valenciana, los dominios son los siguientes:
- Catalánides y Maestrat: Estos dos dominios son similares. En el Cretácico inferior aparece sedimentación asociada a zonas de fracturación. Según se avanza por el período, aparecen terrígenos, calizas, dolomías y, de nuevo, terrígenos.
- Ibérica Suroccidental: Dentro de sus límites se encuentra el Macizo Valenciano.
- Prebético: En la zona externa aparecen terrígenos del Cretácico inferior. Hacia final del período se produce un hundimiento de la zona interna, mientras que la externa emerge.
- Subbética: Los sedimentos están mal representados.
En el Pirineo catalán, se ha establecido correlación entre el Cretácico inferior del anticlinal de Bóixols - Muntanya de Nargó y la Roca de Narieda. Al sur del primero aparecen varios episodios de compresión a partir del Campaniense.[8]
Norteamérica

En las Montañas Rocosas (Estados Unidos), los sedimentos cretácicos se presentan de manera alterna con origen marino o terrestre. A lo largo del margen occidental casi ninguno de los sedimentos que posee es marino.[9] En las Montañas Talkeetna (Alaska), los estratos cretácicos tampoco son marinos.[10]
Las sierras del suroeste del estado de Nuevo León (México) se constituyen a partir de rocas marinas que van desde el Jurásico superior hasta el Cretácico superior.[11] En esta zona, abundan los terrígenos del Cretácico superior. La secuencia estratigráfica de la parte sur del Cañón de la Boca es:
- Calizas del Aptiense.
- Calizas del Albiense y un paleoambiente de talud superior.
- Calizas y lutitas alternadas del Cenomaniense y un paleoambiente de talud.
- Calizas arcillosas y lutitas del Turoniense y paleoambiente de cuenca.
- Lutitas y calizas del Coniaciense y paleoambiente de cuenca.[12]
Sudamérica
En la zona más meridional de América del Sur, se produjeron grandes eventos tectónicos, en parte debido a una compresión.[13][14][15] Una localización bien conocida es la bahía Thetis, en los Andes argentinos. Aquí se reconocen tres formaciones:
- Bahía Thetis: Fangolitas, turbiditas arenosas y conglomerados del Maastrichtiense. Existen una gran cantidad de ammonites y foraminíferos.
- Policarpo: Fangolitas del Maastrichtiense-Daniense.
- Tres Amigos: Conglomerados, areniscas y fangolitas del Paleoceno.[16]
Paleogeografía
Durante el Cretácico, el nivel de los mares estaba en continuo ascenso. Este crecimiento llevó al nivel del mar hasta cotas jamás alcanzadas anteriormente, incluso zonas anteriormente desérticas se convirtieron en llanuras inundadas. En su punto máximo, solamente un 18 % de la superficie de la Tierra estaba sobre el nivel de las aguas (hoy en día la superficie emergida es del 29 %).
El supercontinente Pangea se fue dividiendo durante el Mesozoico para dar lugar a los continentes actuales, aunque con posiciones sustancialmente diferentes. A principios del Cretácico existían dos supercontinentes: Laurasia y Gondwana, separados por el mar de Tetis. A finales del Cretácico los continentes comienzan a adquirir formas semejantes a las actuales. La progresiva separación de los continentes (o de las placas tectónicas por la deriva continental) fue acompañada por la formación de amplias plataformas y arrecifes.
El sistema de fallas del Jurásico inferior había separado Europa, África y el continente norteamericano, aunque estas masas permanecieron próximas entre sí.[17] La India y Madagascar se estaban alejando de la costa oriental africana. En la India se produjo un episodio de vulcanismo masivo entre finales del Cretácico y principios del Paleoceno. La Antártida y Australia, todavía juntas, se alejaron de Sudamérica y derivaron hacia el este. Estos movimientos crearon nuevas vías marinas, entre ellas los primitivos Atlántico septentrional y meridional, así como el mar Caribe y el océano Índico.

Mientras el Atlántico se ampliaba, las orogenias que habían empezado durante el Jurásico continuaron en la cordillera de Norteamérica, mientras que la Orogenia Nevada fue seguida por otras orogenias como la Orogenia Laramide. Una importante masa de agua se extendía desde las aguas del Polo Norte hasta la península de Yucatán y México. Otra vía marina cruzó África a través de la región del Sahara central. El mar de Tetis, que anteriormente limitaba con el sur de Europa, creció hasta cubrir las islas británicas, Europa central, el sur de Escandinavia y la Rusia europea.
El efecto de todo ello fue la división de la Tierra en doce o más masas de tierra aisladas, lo cual favoreció el desarrollo de faunas y floras endémicas. Estas poblaciones producto de su aislamiento en los continentes insulares del Cretácico superior, evolucionaron hasta generar gran parte de la actual diversidad de la vida terrestre actual. En las regiones cretácicas de latitudes superiores a los 50º tanto meridionales como septentrionales se originaron enormes yacimientos de carbón. En el intervalo comprendido entre hace 120 y 75 millones de años, el mar de Tetis rebosaba de microplancton que se convirtió en petróleo (más de la mitad de las reservas petrolíferas mundiales conocidas corresponden a yacimientos originados en Tetis, como el golfo Pérsico, el norte de África, el golfo de México y Venezuela). También destacó la fragmentación y destrucción de conchas y rocas en la evolución de la bioerosión.
Paleoclimatología

Las temperaturas ascendieron hasta alcanzar su máximo punto hace unos 100 millones de años, en los cuales no había prácticamente hielo en los polos. Los sedimentos muestran que las temperaturas en la superficie del océano tropical debieron haber sido entre 9 y 12 °C más cálidas que en la actualidad, mientras que en las profundidades oceánicas las temperaturas debieron ser incluso 15 o 20 °C mayores.[18] En realidad el planeta no debió de ser mucho más cálido que en el Triásico o el Jurásico, pero el gradiente de temperatura entre los polos y el ecuador debió de ser más suave; esto produjo que las corrientes de aire del planeta amainaran, contribuyendo a reducir las corrientes oceánicas y por tanto a océanos más estancados que hoy, evidenciados por extensas deposiciones de pizarra.[19] Después del Cretácico medio las temperaturas iniciaron un lento descenso que fue acelerándose progresivamente y, en los últimos millones de años del período, la media de las temperaturas anuales del oeste norteamericano había disminuido desde los 20 °C hasta los 10 °C.
Paleozoología
Durante el Cretácico, varias clases de reptiles llegaron a su apogeo, especialmente los dinosaurios, que habitaron por aquel entonces todas las regiones del planeta. Al finalizar el período la mayoría de la fauna, tanto terrestre como marina, sufrió la extinción masiva del K-T.
Paleozoología marina

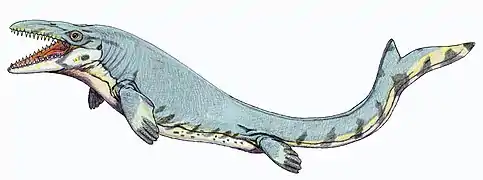
Al final del Cretácico, el plancton oceánico había evolucionado hasta adquirir un carácter completamente moderno. Las diatomeas originadas en el Jurásico e incluso mucho antes, experimentaron su gran expansión en el Cretácico medio, junto con dinoflagelados. Lo mismo ocurre en los mares cálidos con el nanoplancton calcáreo y los foraminíferos planctónicos (globigerinas), que habían aparecido en el Jurásico. A partir de este momento el nanoplancton ha contribuido en cantidades inmensas a la formación de los sedimentos calcáreos conocidos con el nombre de creta. Por encima del lecho nadaban los ammonites, belemnites y peces de varios tipos con una nueva generación de reptiles acuáticos. Los ictiosaurios habían desaparecido prácticamente a finales del Cretácico inferior,[20] y su lugar fue ocupado por nuevos tiburones de gran tamaño y por grandes teleósteos como el Xiphactinus, que medía entre 2 y 4 metros de longitud. Estos a su vez coexistían con grandes tortugas marinas como Archelon, de más de 3 metros de longitud y con unas aletas que, extendidas ambas, superaban la longitud del animal. Eran también comunes las rayas. Entre los reptiles, los elasmosáuridos, un grupo de plesiosaurios de cuello largo, alcanzaban longitudes de hasta 12 metros. Los mosasaurios, que alcanzaban hasta longitudes de 17 metros, eran considerados los más feroces depredadores marinos en el Cretácico superior. Se trataba de grandes lagartos (los mayores que han existido) emparentados con las actuales serpientes (aunque durante mucho tiempo se los ha relacionado con los varanos). Los mosasaurios poseían largas mandíbulas con dientes afilados, juntas a un cuerpo delgado y extenso con extremidades en forma de paleta. Debían alimentarse de peces, aunque se han hallado fósiles de ammonites con marcas de hileras dentales que encajan exactamente con el modelo de dentición de ciertos mosasaurios.
 Jeletzkytes (Ammonoidea)
Jeletzkytes (Ammonoidea) Belemnitella (Belemnoidea)
Belemnitella (Belemnoidea)




En cuanto al bentos, comenzó a ofrecer un aspecto moderno, y continuó la decadencia de los braquiópodos. Asteroidea y Hexacorallia siguieron su diversificación y los foraminíferos bentónicos ya tenían una diversidad similar a la actual. Dentro de Bryozoa se produce una gran expansión de los queilostomados incrustantes, con más de 100 géneros. Los neogasterópodos se difundieron con rapidez, y casi todos sus miembros estaban dotados de nuevas capacidades predatorias (la de perforar conchas y succionar la carne contenida en ellas, la de emponzoñar a sus víctimas, o tragarse enteros a peces pequeños). Otros predadores eran los crustáceos, representados en el Jurásico por criaturas semejantes a la langosta y que en el Cretácico están constituidos por verdaderos cangrejos y langostas (abrían las conchas forzándolas con sus pinzas, como los actuales). Los bivalvos se enterraban cada vez más profundamente en el sedimento para escapar de predadores, o bien desarrollaron conchas muy macizas o espinas para disuadirlos. Un grupo de bivalvos, los inocerámidos, poseía conchas de casi 1,8 metros de longitud. Durante un tiempo, la capacidad de los corales para construir arrecifes fue prácticamente igualada por los rudistas (bivalvos ostreidos con conchas cónicas de hasta 1 m de altura). Se fijaban a una superficie dura del fondo aglutinándose entre ellos y llegando a formar inmensos arrecifes en todos los mares del mundo. Con toda probabilidad estas densas construcciones les protegían de ser devorados.
Paleozoología aérea
Los pterosaurios cretácicos desarrollaron gran variedad de formas que siguieron unas determinadas tendencias.
- Tamaño en aumento a lo largo del Cretácico.
- Pérdida progresiva de los dientes.
- Ahuecamiento de los huesos de las partes no sometidas a los principales esfuerzos.
Destaca especialmente Pteranodon. Las aves fósiles en el Cretácico son raras, excepción hecha de descubrimientos aislados procedentes del Cretácico inferior de un pájaro chino descrito en 1992, y de los españoles (1988-1992) Iberomesornis[21] y Concornis del Cretácico inferior de Cuenca.




Los mamíferos todavía eran un componente pequeño y relativamente menor de la fauna. Por supuesto, los reptiles arcosaurios, sobre todo los dinosaurios tuvieron un lugar muy importante en el Cretácico. De hecho, fue el periodo en el que mayor auge y desarrollo alcanzaron. Los principales yacimientos de dinosaurios se encuentran en las Facies Wealden (Cretácico inferior) y en la formación Nemegt del Cretácico Superior en Mongolia, en la homónima cuenca de Nemegt, en el desierto del Gobi. El Wealden incluye amplias extensiones de sedimentos continentales y lacustres que se extienden por el sureste de Inglaterra y de la isla de Wight aunque también está ampliamente representado en el norte de España. En el plano ambiental era una región dominada por helechos, atravesada por cursos de agua en cuyas riberas crecían cícadas y que desembocaban en pantanos y lagunas. Los dinosaurios dominantes en la zona eran los ornitópodos, un grupo surgido a finales del Jurásico. También habría saurópodos, aunque más escasos. En la península ibérica son abundantes los yacimientos de icnitas siendo mucho menos importantes los restos de dinosaurios aunque recientemente están apareciendo numerosos restos (Teruel, Valencia, La Rioja). En La Rioja aparece uno de los mayores yacimientos del mundo de huellas tanto de terópodos carnívoros como ornitópodos herbívoros.
Los dinosaurios del Cretácico
Durante esta época los dinosaurios alcanzaron una gran radiación adaptativa. Es por ello que existían una gran cantidad de especies con modos de vida y morfologías muy dispares.
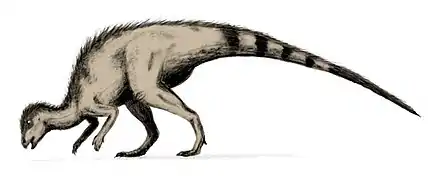
- Ornitópodos: La gran difusión de los ornitópodos a lo largo del Cretácico, así como su diversidad entre las distintas faunas autóctonas, constituyen indicios del éxito que alcanzaron. En el Cretácico inferior nos encontramos con Iguanodon e Hypsilophodon. Iguanodon era el más común de los grandes ornitópodos. Era un bípedo (aunque ha existido cierta controversia histórica a este respecto) de 10 metros de longitud, con el cráneo similar al de un caballo, con largas mandíbulas y ojos en posición muy posterior. Sus mandíbulas estaban armadas con seres de dientes reemplazables, lo cual mantenía joven su dentición trituradora. La zona anterior de la boca estaba desprovista de dientes y presentaba una estructura ósea, instrumento de precisión para cortar hojas y otra vegetación. El verdadero progreso radicaba en su capacidad de masticar. En general caminaba sobre dos patas, lo que le permitía alcanzar las capas altas de los árboles, pero también podía desplazarse sobre cuatro patas, a juzgar por la morfología de sus manos (las garras son en realidad pequeñas pezuñas). La uña del pulgar, en cambio, es una larga y afilada púa,[22] tan distinta de las demás que cuando fue hallada, en el siglo pasado, se identificó como un cuerno nasal. Quizá se trataba de un arma de utilidad contra los depredadores. Hypsilophodon es el segundo dinosaurio más frecuente del Wealden. Era un pequeño animal (de 3 a 5 metros de longitud) con piernas diseñadas para correr velozmente y una rígida cola, que le equilibraba durante la carrera.
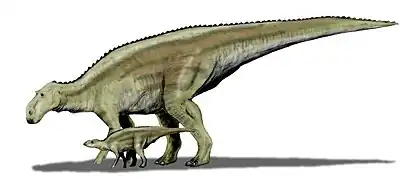
- Los hadrosaurios eran grandes, de 10 a 15 metros de longitud, y todos poseían cuerpos que básicamente se ajustaban al modelo de los iguanodóntidos. La parte anterior del hocico es aplanada, con la típica forma de pico de pato, y los dientes están dispuestos en múltiples hileras, alcanzando a menudo los 500 dientes por segmento mandibular. Muchos hadrosaurios presentaban proyecciones de los huesos nasales y premaxilares del cráneo que, dirigidas hacia atrás, se traducían en una amplia variedad de crestas (algunas en forma de placa, otras de tubo, otras de púa). En los hadrosaurios, el aire de los conductos nasales y premaxilares discurría por el interior de la cresta, que a menudo sufría torsiones o giros. La función de estas crestas era proporcionar indicaciones tanto visuales como sonoras. Destacaron Maiasaura y Parasaurolophus.
- Los ceratópsidos vivieron en Norteamérica y Asia, y tenían una cabeza grande y picuda, que vista desde arriba tenía forma triangular, presentaban un número variable de cuernos y un escudo óseo que protegía la región de los hombros y que también constituía el plano de inserción de la poderosa musculatura mandibular. Los mayores ceratópsidos alcanzaron los 9 metros de longitud, con cuernos de casi 1 metros y pesos de hasta 6 toneladas como Triceratops, hallado en el oeste de Norteamérica. También destacó Protoceratops.
- Los paquicefalosaurios eran otro grupo de ornitópodos bípedos con gruesos cráneos. Están muy extendidos en el Cretácico superior de Norteamérica y Asia. Stegoceras sólo medía 2 metros de longitud, aunque su bóveda craneana era excepcionalmente gruesa (25 centímetros, casi la mitad de la longitud del cráneo). Se ha sugerido que los paquicefalosaurios luchaban entre sí por el apareamiento de una forma parecida a la de los carneros y otros herbívoros semejantes. También destacó Pachycephalosaurus.


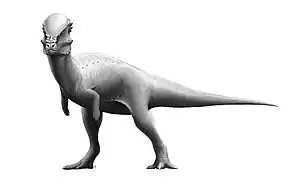
- En el Cretácico tardío se diversificaron los anquilosaurios, que habían aparecido en el Jurásico. Estaban estrechamente relacionados con los estegosaurios, y eran con frecuencia animales de gran tamaño, de hasta 10 metros de longitud y 6 toneladas de peso. Se especializaron en el blindaje, con presencia de placas ódeas bajo la piel del dorso, cuello, cola y flancos, así como de refuerzos óseos en el cráneo, e incluso de «postigos» óseos por encima de los ojos. Poseían pequeños cuernos óseos en la cabeza y variadas pautas de espinas, también óseas, dispuestas sobre el dorso. Los anquilosaurios más tardíos también poseían una masa de hueso fusionado en el extremo de la cola que debía ejercer la función de una pesada maza oscilante, que podía golpear las piernas de un atacante mientras el animal se mantenía pegado al suelo.
- Terópodos: Entre los terópodos carnívoros podríamos incluir diversas formas. En 1983, el descubrimiento de Baryonyx llevó al reconocimiento de un nuevo grupo de carnívoros en este animal, conocido por un solo esqueleto. El rasgo más destacado es su inmensa garra en forma de hoz. La garra de Baryonyx medía 30 centímetros, más que considerable para un animal de 9 metros. Curiosamente, aún no está claro si la garra estaba unida a la mano o al pie. En la región de su caja torácica, fueron halladas escamas de pez, de lo que se deduce que era piscívoro. El desarrollo de garras curvas, especialmente diseñadas para destripar a las presas, parece haberse dado de forma independiente en un cierto número de terópodos. Tyrannosaurus rex (Norteamérica) y el mongol Tarbosaurus, muy semejante al anterior, eran probablemente los máximos depredadores.[23] Tyrannosaurus fue el mayor carnívoro terrestre de su tiempo, solo ligeramente superado por Giganotosaurus y Spinosaurus. El tiranosaurio alcanzaba una longitud de trece metros, una altura de 4,8 metros y un peso de 5 toneladas. En sus mandíbulas se alineaban dientes de 15 centímetros de longitud. Las patas traseras de Tyrannosaurus eran muy fuertes y macizas, con tres dedos en cada pie. Sus brazos, en cambio, parecen casi ridículos: extremadamente delgados y tan cortos que no llegaban a la boca, y con solo dos dedos. La sólida cabeza no presenta el sistema de ahorro de peso observado en otros carnívoros, y parece estar diseñada para golpear. Un depredador tan grande no podía correr velozmente durante mucho tiempo. Seguramente se alimentaba de grandes herbívoros a los que tendía emboscadas, o bien de animales ya debilitados o muertos.






- Otros grandes carnívoros fueron Carnotaurus y Spinosaurus. También existieron terópodos depredadores de menor talla (Velociraptor, Deinonychus, Oviraptor, Gallimimus, Avimimus). Al parecer, todos los dinosaurios del clado Maniraptora y quizás algún otro grupo de Theropoda tenían plumas o protoplumas. Las aves fueron los únicos dinosaurios que sobrevivieron al final del Cretácico.
Paleobotánica
Pocos cambios han afectado de forma tan profunda al paisaje y a la ecología de la Tierra como la llegada de las angiospermas. Sin embargo, durante el Cretácico inferior, las gimnospermas seguían dominando. El único grupo que seguía prosperando era el de las coníferas. Las cícadas y los ginkgos estaban ya en decadencia y las bennettitales afrontaban su inminente extinción. Las angiospermas aparecieron en algún momento del Cretácico inferior, iniciando una progresiva difusión y diversificación que las ha conducido hasta sus actuales 250 000 especies, en contraste con las 550 especies de coníferas existentes. A diferencia de las gimnospermas, portadoras de semillas desnudas, las angiospermas encierran sus semillas en el interior de un ovario, que las protege de las infecciones fúngicas, de la desecación y del ataque de los insectos. Los fósiles de angiospermas más antiguos datan del Cretácico inferior, hace unos 130-120 millones de años: Consisten en granos de polen llamados Clavatipollenites, procedentes de Wealden, al sureste de Inglaterra. El polen y las hojas se hicieron más frecuentes hace unos 120-100 millones de años. En el este de Norteamérica, Rusia e Israel, han sido extraídas varias flores y frutos de angiospermas primitivas muy bien conservadas, relacionadas aparentemente con las modernas magnolias y sicomoros.

Durante el Cretácico superior (en el Campaniense) habían hecho su aparición unas 50 de las familias modernas (de un total de 500), entre las cuales se contaban las hayas, higueras, abedules, acebos, magnolias, robles, palmeras, sicomoros, nogales y sauces. El polen y las hojas son los fósiles más comunes. Los niveles de polen angiospérmico aumentaron desde el 1 % del contenido total de polen hasta 40 % tan solo en 20 millones de años, lo que demuestra el desenfrenado crecimiento de las plantas con flores. Las hojas inicialmente eran pequeñas de nervadura irregular y bordes lisos y sencillos. En el Cretácico superior eran más grandes y con el margen serrado o lobulado, y las nervaduras presentaban patrones mucho más regulares. Los primeros hábitats de las angiospermas parecen haber sido áreas fuertemente perturbadas por corrientes o inundaciones. Los matorrales, malas hierbas con rápido crecimiento y pequeños arbustos, pudieron dominar estos medios, si eran capaces de producir semillas con rapidez. Las coníferas tienen el grave problema de que, después de la fertilización, tienen que esperar el paso del invierno antes de liberar sus semillas por lo que necesitan dos años para germinar. Tal vez el paso de dinosaurios migratorios, con el consiguiente pisoteo y fertilización a que se sometían al suelo, favoreciera el desarrollo oportunista de hierbas y matorrales.
Interacción dinosaurio-planta
Las plantas jurásicas habían desarrollado sistemas de defensa contra los dinosaurios ramoneadores que devoraban la vegetación de las capas altas (saurópodos y estegosaurios). Las gimnospermas desarrollaron espinas, venenos o sabores desagradables para protegerse de la destrucción de sus capas altas por la abundancia de ramoneadores, aunque sus vástagos, a ras del suelo, no necesitaban tal protección. La extinción de los ramoneadores de capas altas y la aparición de eficientes especies devoradores de plantas bajas, propició la evolución de plantas capaces de crecer y generar semillas con rapidez para perpetuar sus especies. Las angiospermas tenían esta capacidad y las gimnospermas no; por tanto, los nuevos dinosaurios pacedores habían provocado la aparición de las angiospermas. Otros expertos contemplan este proceso justo a la inversa, es decir, fue la difusión de las angiospermas la que habría favorecido la aparición de herbívoros pacedores especialistas de capas bajas a expensas de los ramoneadores de capas altas. Todavía no se ha aclarado cuál de las dos teorías se aproxima más a la verdad.
Primeras flores

Las plantas con flores no aparecen hasta el Cretácico, y las primeras no tenían pétalos. Su desarrollo seguramente fue posible por su asociación con los insectos. Se puede decir que insectos y plantas coevolucionaron. Es probable que los escarabajos polinizaran algunos vegetales considerados próximos al ancestro de las angiospermas, como las bennettitales, cuyos órganos reproductores, semejantes a una flor, podían atraer a los polinizadores mediante perfumes o pálidos colores, «entrenando» así a ciertos insectos en la respuesta a este tipo de señuelos. Otros insectos polinizadores del Jurásico y Cretácico temprano pudieron pertenecer a los dípteros, himenópteros o ser incluso pequeñas mariposas nocturnas. Algunos de los insectos mejor conocidos del Cretácico inferior provienen del Wealden del sureste de Inglaterra e incluyen libélulas, cucarachas, grillos, chinches, escarabajos, moscas, avispas y termitas entre otros. Algunos himenópteros tienen granos de polen en su interior, lo que prueba que aquel formaba parte de su dieta y los pelos especializados de las patas de las avispas esfécidas, surgidas en el Cretácico inferior, demuestran actividad polinizadora. También se han encontrado insectos excepcionalmente conservados en el Cretácico inferior de Baissa (Siberia).
La primera abeja cretácica no fue descubierta hasta 2006 por George Poinar, hijo, en ámbar procedente de Birmania. Se trata de Melittosphex burmensis. Presenta una combinación de caracteres de abeja y de avispa.
El ámbar procedente de Nueva Jersey es uno de los yacimientos más importantes pertenece al Cretácico inferior de Álava y ha proporcionado desde 1996 más de 1500 fósiles de artrópodos fundamentalmente insectos (55 % dípteros, 24 % himenópteros que incluyen la abeja Cretotrigona prisca). Las primeras angiospermas seguramente eran polinizadas por diversos insectos. Las relaciones más selectivas se desarrollaron posiblemente a partir del Cretácico superior con la aparición de las avispas vespoideas, actualmente polinizadoras de pequeñas flores de simetría radial. Al final del Cretácico y durante el Terciario, las flores fueron adaptándose más y más a un solo tipo de insecto polinizador, abejas incluidas. La aparición de termitas en el Cretácico inferior y la de abejas y hormigas en el superior, indica avances cruciales en la conducta social de los insectos. El origen de estos insectos sociales, de gran éxito en la actualidad, podría estar relacionado con la radiación de las angiospermas.
Extinción en masa

A este acontecimiento del final de Cretácico se le conoce como episodio K–T (o K/T), del alemán Kreide/Tertiär para Cretácico/Terciario, actualmente K–Pg (o K/Pg) de Cretácico-Paleógeno, una vez que el Terciario ha sido descartado formalmente por la Comisión Internacional de Estratigrafía. La extinción masiva de finales del período Cretácico exterminó a los dinosaurios, pterosaurios, reptiles nadadores, plesiosaurios y mosasaurios, Ammonoidea, rudistas e inocerámidos. El nannoplancton calcáreo y los foraminíferos planctónicos experimentaron pérdidas importantes pero se recuperaron durante la Era Cenozoica. Sin embargo, no fue mayor que otras extinciones masivas e incluso fue mucho menos catastrófica que la extinción del Pérmico tardío. Algunos de los grupos que se extinguen muestran claras pautas de reducción progresiva de la diversidad durante los últimos 10 millones de años del Cretácico, mientras que otros parecen desvanecerse completamente justo en el momento de la transición. También hay otros grupos, como los ictiosaurios, supuestamente extinguidos en el límite K-Pg, pero que realmente habían partido mucho antes. Entre los supervivientes se hallan la mayor parte de plantas y animales terrestres (insectos, caracoles, ranas, salamandras, tortugas, lagartos, serpientes, cocodrilos y mamíferos placentarios) y la mayoría de invertebrados marinos (estrellas de mar y Echinoidea, moluscos, artrópodos) y de peces. Durante el Albiense se produjo una intensa extinción de insectos.

Entre las causas se encuentran:
- Hipótesis climáticas: Solo resultaron afectadas las faunas tropicales, con extinción de los rudistas y otros pobladores del mar de Tetis, mientras que las faunas de latitudes elevadas permanecieron intactas. Una posible causa del enfriamiento la hallamos en los movimientos tectónicos que estaban separando Australia de la Antártida. Las frías corrientes profundas del océano meridional habrían sido encauzadas hacia las aguas ecuatoriales, más cálidas, del mar de Tetis. Las aguas más frías, combinadas con el descenso del nivel del mar, habrían afectado las temperaturas ecuatoriales, y desprovisto al clima de la influencia moderadora de los mares cálidos. Los cambios climáticos resultantes de ello generarían condiciones globales más frías, y climas muy extremos en los interiores continentales.

- Hipótesis extraterrestres: Las hipótesis más extrañas provienen de este grupo. Si bien son muy numerosas, destaca sobremanera una posible disminución de la radiación solar que afectó toda la cadena trófica. Otra de estas teorías argumenta que una supernova relativamente cercana pudo aumentar el nivel de radiación en la Tierra y afectar a todos los organismos terrestres y acuáticos de la zona fótica.
- Impacto meteorítico: Luis Álvarez y Walter Álvarez, a partir de una capa de iridio (formada por el impacto de un meteorito), esferulitos vítreos y cuarzos fracturados, postularon que apareció una nube de polvo oscureciendo el globo.[24] Posteriormente se repitieron fenómenos de lluvia ácida, y desapareció el fitoplancton. Las críticas que se hacen a esta teoría es que las desapariciones no fueron súbitas. Muchas especies atravesaron el límite sin pérdidas. El fitoplancton calcáreo desaparece cerca de la capa de iridio, a unos 10 000 años. Ammonoidea desaparece gradualmente de 10 a ninguno en 2 millones de años. Unos 12 millones de años por debajo de la capa de iridio está el último. Los inocerámidos desaparecen 60 000 años antes de la capa de iridio.
El balance sobre la extinción K–Pg indica de momento un empate entre el modelo catastrófico basado en un impacto y el modelo de enfriamiento global progresivo. Existen pruebas de ambos y ambos pueden tener importancia. Un tercer conjunto de causas reside en el interior de las plantas y animales mesozoicos: no se sabe cuáles fueron los rasgos biológicos que determinaron la muerte de unos y la supervivencia de otros. Todas estas líneas de investigación y otras convergen en el episodio K–Pg y forman un gran debate.
Véase también
Notas y referencias
- Los colores corresponden a los códigos RGB aprobados por la Comisión Internacional de Estratigrafía. Disponible en el sitio de la International Commision on Stratigraphy, en «Standard Color Codes for the Geological Time Scale».
- «International Commission on Stratigraphy: International Chronostratigraphic Chart v2020/01» (en inglés). enero de 2020. Consultado el 5 de mayo de 2020.
- (1972) Glossary of Geology, 3rd ed., Washington, D.C.: American Geological Institute, p. 165.
- (1974) Great Soviet Encyclopedia, 3rd ed. (in Russian), Moscow: Sovetskaya Enciklopediya, vol. 16, p. 50.
- «International Commission on Stratigraphy: International Chronostratigraphic Chart» (en inglés). octubre de 2021. Consultado el 28 de diciembre de 2021.
- Cohen, K. M., Finney, S. C., Gibbard, P. L. & Fan, J.-X. (2013; actualizado 2022). «The ICS International Chronostratigraphic Chart». Episodes 36. pp. 199-204. Consultado el 30 de diciembre de 2022.
- Universidad de Valencia. «Estratigrafía del Cretácico». Consultado el 27 de enero de 2008.
- Ullastre, J. & Masriera, A. 2006. El anticlinal de Bóixols - Muntanya de Nargó: consideraciones estratigráficas y estructurales basadas en una nueva cartografía geológica (Pirineo catalán, España). Treb. Mus. Geol. Barcelona, 14: 5-35
- Weimer, R.J. 1960. Upper Cretaceous stratigraphy, Rocky Mountain area. AAPG Bulletin; v. 44; no. 1; p. 1-20
- Brian A. Hampton. «Regional Upper Triassic-Cretaceous stratigraphy along the Northern margin of Wrangellia, Southern Alaska: Implications for a shared history of basin development in the Alaska range suture zone». Archivado desde el original el 13 de febrero de 2009. Consultado el 28 de enero de 2008.
- Padilla y Sánchez, R. J. 1978. Geología y estratigrafía (Cretácico superior) del límite suroeste del estado de Nuevo León. Univ. Nal. Autón. México, Inst. Geología. (2)1: 37-44
- Ángeles-Villeda, M.A., Hinojosa-Espinosa, J.J.,López-Oliva, J.G., Valdés-González, A. & Livas-Vera, M. 2005. Estratigrafía y microfacies de la parte sur del Cañón La Boca, Santiago, Nuevo León, México. Revista Mexicana de Ciencias Geológicas, v. 22, núm. 2, 2005, p. 272-281
- Suárez, M.; De La Cruz, R.; Bell, C.M. 2000. Timing and origin of deformation along the Patagonian fold and thrust belt. Geological Magazine, Vol. 134, p. 345-353.
- Biddle, K.T.; Uliana, M.A.; Mitchum, R.M. Jr.; Fitzgerald, M.G.; Wright, R.C. 1986. The stratigraphic and structural evolution of the central and eastern Magallanes Basin, southern South America. In Foreland Basins (Allen, P.A.; Homewood, P.; editors). International Association of Sedimentologists, Special Publication No. 8, p. 41-61.
- Dalziel, I.W.D.; De Wit; M.J.; Palmer, K.F. 1974. Fossil marginal basin in the southern Andes. Nature, No. 250, p. 291-294
- Olivero, E.B., Malumián, N., Palamarczuk, S. 2003. Estratigrafía del Cretácico Superior-Paleoceno del área de Bahía Thetis, Andes fueguinos, Argentina: acontecimientos tectónicos y paleobiológicos. Rev. geol. Chile, vol.30, no.2, p.245-263. ISSN 0716-0208.
- Canudo, J. I. 2006. La ambigüedad paleobiogeográfica de los dinosaurios ibéricos durante el Cretácico Inferior. En (Colectivo Arqueológico-Paleontológico Salense, Ed.): Actas de las III Jornadas sobre Dinosaurios y su Entorno. 21-45. Salas de los Infantes, Burgos, España.
- "Warmer than a Hot Tub: Atlantic Ocean Temperatures Much Higher in the Past" PhysOrg.com. Retrieved 12/3/06.
- Stanley, pp. 481-2
- Smithsonian Natural Museum of Natural History. «Cretaceous:Life in the Cretaceous Seas». Consultado el 31 de diciembre de 2007.
- Museo de las Ciencias de Castilla-La Mancha. «Museo de las Ciencias». Archivado desde el original el 11 de noviembre de 2007. Consultado el 31 de diciembre de 2007.
- Senckenberg, forschungsinstitut und naturmuseum. «Iguanodon». Archivado desde el original el 13 de febrero de 2009. Consultado el 31 de diciembre de 2007.
- Universidad de Berkeley. «The Tyrannosauridae». Consultado el 31 de diciembre de 2007.
- «La extinción de los dinosaurios. El límite Cretácico-Terciario». Archivado desde el original el 5 de enero de 2008. Consultado el 1 de enero de 2008.
Bibliografía
- Allain, R. (2001). «Redescription de Streotospondylus altdorfensis, le dinosaure théropode de Cuvier, du Jurassi-que de Normandie». Geodiversitas, 23(3): 349-367
- Álvarez, W. (1998). Tyrannosaurus rex y el Cráter de la Muerte. Crítica, Drakontos, Barcelona, España, 201 p.
- Amo Sanjuán, O., Canudo, J. I. y Cuenca-Bescós, G. (2000): «First record of elongatoolithid eggshells from the Lower Barremian (Lower Cretaceous) of Europe (Cuesta Corrales 2, Galve Basin, Teruel, Spain)». En: First International symposium on Dinosaur eggs and babies. Extendend abstracts: 7-14.
- Benton (1995). Paleontología y evolución de los vertebrados. Lleida: Editorial Perfils. 369 págs. ISBN 84-87695-16-7
- Dressler, B.O.; Grieve, R.A.F. y Sharpton, V.L. (Eds.) (1994). Large Meteorite Impacts and Planetary Evolution. Geological Society of America, Special Paper, 293 ISBN 0-8137-2293-4
- Mac Leod, Norman y Keller, Gerta (Eds.) (1994). Cretaceous-Tertiary Mass Extinctions Biotic and Environmental Changes. [Prefacio de Niles Eldredge]
- Skinner, Brian J. y Porter, Stephen C. (1995). The Dynamic Earth: An Introduction to Physical Geology (3ª ed.). New York: John Wiley & Sons, Inc. 557 págs. ISBN 0-471-59549-7
- Taylor, P.D. y Wilson, M.A. (2003). «Palaeoecology and evolution of marine hard substrate communities». Earth-Science Reviews, 62: 1-103.
- Upchurch, P., Hunn, C. A. y Norman, D. B. (2002): «An analysis of dinosaurian biogeography: evidence for the existence of vicariance and dispersal patterns caused by geological events». Proceedings of the Royal Society of London, 269: 613-621.
Enlaces externos
 Wikimedia Commons alberga una galería multimedia sobre Cretácico.
Wikimedia Commons alberga una galería multimedia sobre Cretácico. Wikcionario tiene definiciones y otra información sobre Cretácico.
Wikcionario tiene definiciones y otra información sobre Cretácico.- Cretácico La deriva continental (Proyecto Celestia)
- http://www.duiops.net/dinos/cretacico.html
- UCMP Berkeley Cretaceous page
- https://web.archive.org/web/20030919005846/http://homepage.mac.com/uriarte/cretacico.html
- https://www.webcitation.org/query?id=1256591525442145&url=www.geocities.com/jmb_2.geo/cretaci_e.html
 Datos: Q44626
Datos: Q44626- Multimedia: Cretaceous / Q44626