Evento de extinción del Cuaternario
El evento de extinción del Cuaternario tardío, Extinción de Megafauna del Cuaternario —también citado en la literatura científica como LQE (Late Quaternary Extinctions)[1] o QME (Quaternary Megafauna Extinction)[2] por sus siglas en inglés— abarca el conjunto de extinciones masivas de megafauna que viene sufriendo el planeta durante el Cuaternario tardío, en un lapso de tiempo geológico extremadamente corto, que se extiende desde hace unos 50.000 años hasta la actualidad (se estima que la vida apareció en la Tierra hace al menos 3.700 millones de años).[3][4][5]

En este evento de extinción masiva se han perdido 2/3 de todos los géneros de grandes mamíferos del planeta y la mitad de su megafauna (considerada como animales con un peso mayor de 44 kilos).[6][7] Entre las posibles causas que desarrollaron como hipótesis los paleontólogos y los ecólogos durante el SXX están la caza excesiva por el Homo sapiens y los cambios climáticos naturales. La hipótesis del cambio climático fue la predominante durante el SXX, mientras que la de la caza excesiva va ganando partidarios desde que se enuncia por primera vez en los 60,[8] y a partir del SXXI comienza a ser defendida ya de forma categórica en numerosos artículos científicos, a medida que se va estudiando el evento de forma global, y los análisis comparativos mediante modelos estadísticos ponen de manifiesto que la relación de causa y efecto más probable con las últimas dataciones de alta precisión, se encuentra entre la expansión del ser humano y las extinciones en masa de megafauna.[1][2][6][7][8][9][10][11][12][13][14][15][16][17][18][19][20][21][22][23][24][25][26][27][28][29][30][31][32][33][34][35][36][37][38][39][40][41]
La ola de extinciones comenzó primero en Asia tropical y Oceanía, hace entre 50.000 y 45.000 años, coincidiendo con la llegada estimada de los primeros hombres modernos, y continuó en la Eurasia holártica con extinciones masivas más dispersas en el tiempo, presentando aparentemente dos pulsos, uno más al sur que tuvo lugar entre 45.000 y 20.000 años atrás, y otra en latitudes septentrionales donde las extinciones en masa llegaron más tarde, de 14.000 a 9.000 años atrás. Este segundo pulso del Paleártico septentrional coincide en el tiempo con la extinción masiva de megafauna de Norteamérica, y ambas son coincidentes a su vez con los primeros registros fósiles de Homo sapiens en Siberia y América del Norte,[6] entonces unidas por el puente de Beringia. Unos 5 siglos después de iniciada la extinción masiva norteamericana, comenzó la extinción en masa de Sudamérica. Desde hace 9 milenios hasta el SXX la extinción masiva de megafauna ha continuado en las islas y archipiélagos de todos los mares, habiéndose comprobado en la mayoría de los casos que la extinción de la megafauna fue muy cercana en el tiempo a la datación de los primeros restos conocidos del hombre moderno en las islas, es el caso por ejemplo de Japón, las Antillas, la Isla de Wrangel, la Isla de Saint Paul, las islas mediterráneas, Madagascar o Nueva Zelanda entre otras.[6][7][42]
Hipótesis sobre la causa de la extinción de megafauna del Cuaternario tardío
Uno de los primeros científicos en llamar la atención sobre la extinción masiva de megafauna del Cuaternario fue Alfred Rusell Wallace, quien en 1876 escribió:
Vivimos en un mundo biológicamente empobrecido, un mundo en el que las formas de vida más grandes, fieras y extrañas han desaparecido. Es desde luego un hecho extraordinario y no lo suficientemente discutido, el de la muerte de tantos grandes mamíferos, no en un solo lugar, sino en más de la mitad de la superficie terrestre.[43]
Hoy se sabe que esa idea de un mundo ecológicamente incompleto apuntada por Wallace en el SXIX —en un momento en que la paleontología era aún una ciencia joven— estaba justificada.[2][7] En los últimos 50.000 años —un instante en la escala del tiempo geológico— el planeta ha perdido una enorme variedad de grandes animales, incluyendo mamutes, mastodontes, rinocerontes lanudos, elefantes europeos, hipopótamos europeos, tigres dientes de sable, perezosos gigantes, gliptodontos, moas, uros, tarpanes, aves elefante y leones marsupiales entre muchos otros. Estas criaturas recientemente perdidas representaban unos 2/3 de los megamamíferos (con un peso de más de 44 kilos): 97 de los 150 géneros de grandes mamíferos han desaparecido entre hace 50.000 y 500 años.[44]

Dada la magnitud de esta extinción masiva de megafauna y sus enormes consecuencias para los ecosistemas terrestres, algunos autores han calificado la extinción de megafauna del Cuaternario de "revolución prehistórica".[29] Estas extinciones no fueron un evento simultáneo, sino un proceso que duró miles de años: la megafauna desapareció en periodos distintos en diferentes partes del planeta.
Desde que Wallace llamó la atención sobre la reciente pérdida masiva de grandes animales, ha habido mucha controversia sobre qué fue lo que causó las extinciones cuaternarias, todas ellas con cuatro elementos comunes: fueron extinciones profundamente selectivas para la megafauna, no hubo reemplazo de las especies extintas —ambas cosas sin precedentes en la historia de la vida—, ocurrieron en presencia de nuestra especie y son inmediatas al momento actual en escala geológica y evolutiva. Se han enunciado diversas hipótesis durante el SXX, de las cuales solo dos fueron asumidas por un número significativo de investigadores como posibles.
Hipótesis del cambio climático
La hipótesis de que la desaparición de la megafauna había sido causada por cambios climáticos fue la predominante durante casi un siglo después de que Wallace llamara la atención sobre el vacío ecológico dejado por la megafauna en la mayor parte del planeta.[43] La hipótesis sigue siendo popular y ha sido propuesta en muchos estudios regionales para explicar la desaparición de la megafauna del Pleistoceno.[45] Sin embargo, cuando se analizan las extinciones desde una perspectiva global, la hipótesis climática es incapaz de explicar los principales patrones del evento de extinción masiva de megafauna del Cuaternario. Así, numerosos estudios están poniendo de relieve en la última década los puntos débiles e incoherencias de esta teoría, destacando los siguientes:

• La hipótesis climática no explica la gran asincronía regional de las extinciones. Los eventos climáticos relacionados con los últimos ciclos glaciales fueron acontecimientos planetarios que afectaron a todo el globo al mismo tiempo: los picos fríos por ejemplo han estado vinculados a períodos secos en regiones tropicales. Sin embargo, las extinciones de megafauna ocurrieron en periodos completamente distintos en lugares muy diferentes del mundo.[6]
• Hubo más de 30 ciclos glaciales a lo largo del Pleistoceno al menos tan intensos como el último, pero sin extinciones masivas asociadas en el registro fósil.[14][46]
• Las extinciones en Oceanía y las islas del planeta no coincidieron con los picos glaciales ni con los cambios de ciclo glacial, sino con periodos estables climáticamente.[47]
• La hipótesis climática implicaría extinciones de numerosas especies de plantas, ya que se ven más afectadas por los cambios climáticos que los animales, pero esta predicción falla ostensiblemente: no ha habido ninguna ola de extinciones florísticas en el Cuaternario tardío.[6]
• Las extinciones presentan una selectividad de tamaño sin precedentes en el registro fósil, ya que afectan casi exclusivamente a especies de megafauna. Sin embargo, los estudios de la última década están revelando que son las especies más pequeñas las más vulnerables y las que se han extinguido en mayor proporción bajo perturbaciones medioambientales provocadas por el clima.[13]

• Los fósiles muestran en el periodo inmediatamente anterior a la extinción una reducción drástica en la edad de madurez sexual. Este fenómeno no se corresponde con lo registrado en extinciones debidas a factores climáticos, pero en cambio sí se ha registrado en poblaciones de grandes mamíferos sujetos a una elevada presión cinegética.[6]
• Por último los cambios climáticos no pueden explicar por qué las extinciones han sido sistemáticamente más recientes en las islas, independientemente de su latitud y longitud, que en los continentes, cuando ante un cambio climático lo esperado es justo lo contrario: las poblaciones insulares, sin posibilidades de emigrar como las continentales, y con un número de ejemplares y un pool genético mucho menores, debieron extinguirse en primer lugar. Además, las extinciones en las islas ocurrieron ya muy avanzado el Holoceno, varios miles de años después del último cambio climático.[8] Así hay numerosos casos, como el del mamut lanudo de Wrangel y Saint Paul, el mamut pigmeo de Santa rosa, los elefantes e hipopótamos de las islas mediterráneas, los perezosos terrestres de las Antillas y la vaca marina de Steller en que las extinciones entre continente e islas estuvieron separadas por varios miles de años de diferencia, y prácticamente en todos los casos parece que coincidieron con la llegada del ser humano.[49][50][51][52][53][28][6]
Aparte de estas incoherencias internas de la hipótesis, hoy se apunta a que procesos de profunda transformación en los ecosistemas que fueron atribuidos originalmente al cambio climático como desencadenantes de la extinción de megafauna —como la expansión de los bosques y los grandes incendios—, probablemente fueron consecuencia y no causa de esa extinción.[11][30][54] Tras la desaparición de los grandes herbívoros, el acúmulo de materia vegetal no consumida y la proliferación de los bosques ante la ausencia de herbívoros habrían llevado primero a un cambio radical de los ecosistemas y después a una recurrencia de grandes incendios, tras los cuales la vegetación mejor adaptada al fuego se habría visto favorecida. Se ha comprobado que esto fue así al menos en el caso de Australia[54][55] Norteamérica[56] Madagascar[57] y varias islas oceánicas. Varios estudios apuntan a que ocurrió algo similar en Sudamérica[58] y otras masas terrestres y biomas afectados por la extinción del Cuaternario, como la estepa del mamut en el Holártico, que era el mayor bioma terrestre hasta su desaparición.[59][31] Algunos autores están señalando que las extinciones de megafauna tuvieron tal impacto en los ecosistemas y el ciclo del carbono que pudieron provocar cambios climáticos.[10]
Sí se apunta a que en la región de Sudamérica, los cambios climáticos podrían haber contribuido a acelerar el proceso de extinción iniciado por la caza insostenible en el caso de algunas especies.[60] En este caso el clima no habría tenido un efecto sinérgico, sino secundario y meramente contributivo.[1][6][61]
Hipótesis de la caza excesiva

A la luz de los estudios estadísticos publicados a partir de 2012, se obtiene una probabilidad de causa-efecto cada vez más sólida entre la llegada de los humanos y las extinciones masivas de la megafauna en cada masa terrestre.[6][11][14][32][1] En la presente década los ecólogos han comenzado a aplicar análisis comparativos basados en modelos matemáticos para dejar atrás el debate de ideas que han protagonizado los partidarios de ambas hipótesis durante el SXX respecto a las causas de la extinción. Estos análisis comparan mediante modelos matemáticos la probabilidad de que una extinción de megafauna a nivel de género o a nivel regional se deba a causas antrópicas, a causas climáticas o a ambas a la vez. Igualmente los análisis de esporas del hongo del estiércol Sporormiella y las catas de polen fósil, n-alcanos y carbón están comenzando a mostrar la secuencia de acontecimientos que siguió a la extinción de megafauna en varios puntos del planeta.[54][55][56]
La crítica de la asociación
La principal crítica a la hipótesis de la caza excesiva que se ha venido esgrimiendo por los partidarios de la hipótesis del cambio climático, es una supuesta baja densidad de yacimientos arqueológicos con evidencias de aprovechamiento de la megafauna, descrita como crítica de la asociación por Meltzer en 1986.[62] Desde entonces esta observación ha sido utilizada por el propio Meltzer y otros autores para concluir que no había habido caza excesiva.[62][41]

Actualmente la alegada ausencia de evidencias de despiece de megafauna se viene corrigiendo a medida que avanzan las excavaciones y los análisis de las colecciones de fósiles. En el caso de Madagascar, una de las extinciones más recientes y sin cambio climático conocido desde la ocupación humana de la isla, los defensores de la hipótesis del cambio climático incidieron durante las últimas décadas del SXX en el hecho de que solo había evidencias firmes de descuartizamiento de aves elefante, aye-ayes extintos e hipopótamos pigmeos, pero no de lémures gigantes. Hoy existen ya varios estudios sobre restos de lémures gigantes que fueron objeto de descuartizamiento, en algunos casos en el periodo inmediatamente posterior a la llegada del hombre.[63][64] Las características de las alteraciones óseas de los huesos de estos lémures gigantes fueron provocadas por herramientas humanas (filos cortantes y marcas de corte cerca de las articulaciones, cortes oblicuos a lo largo de los ejes, fracturas espirales y estriaciones de percusión) e indican que hubo despellejamiento, desarticulación y fileteado de la carne.[63]
Otras regiones en las que la crítica de la asociación alegaba falta de evidencias, están incorporando pruebas muy recientes de cacería humana de megafauna, como en el caso de Australia (noviembre de 2016)[65] y Sudamérica (enero de 2017).[66]

El ecólogo Santos Fernández explicaba en 2016 el caso de los mamutes norteamericanos en relación con la crítica de la asociación: el periodo de convivencia de mamutes y seres humanos, hasta que los mamutes se extinguieron, constituye una parte muy pequeña del amplio rango geológico en que han aparecido restos de mamutes norteamericanos. Cazadores y proboscídeos habrían convivido alrededor de veinte siglos en América del Norte, una mínima fracción del registro geológico con restos de mamutes y restos arqueológicos, y en una localidad concreta la convivencia fue mucho más corta aún.[6] Por lo tanto, es muy complicado encontrar restos arqueológicos que sean precisamente de las pocas décadas o siglos en que los mamutes y los seres humanos coexistieron en una localidad en concreto. En cualquier otro caso, solo se encontrarán mamutes sin seres humanos (antes de la coexistencia) o humanos sin mamutes (después de la coexistencia). El patrón esperado es que una fracción minoritaria de yacimientos fosilíferos de megafauna tendría restos arqueológicos con evidencias de cacería, y eso es lo que se encuentra.[6]
Un artículo de 2002 de Meltzer y otro investigador concluía que el registro arqueológico no apoyaba la hipótesis de la caza excesiva, porque de 76 yacimientos clovis examinados, solo 14 (doce con restos de mamut colombino y dos con restos de mastodonte) presentaban evidencias sólidas de cacería de proboscídeos.[67] A su juicio está proporción era muy pequeña y descartaba totalmente la caza excesiva como motivo de la extinción de proboscídeos en Norteamérica. El artículo fue duramente criticado por emitir juicios de opinión en vez de presentar nuevos análisis comparativos independientes, y por omitir numerosos datos científicos que contradecían las afirmaciones de los autores.[41] Refutando estas conclusiones del artículo de Meltzer, un análisis estadístico comparativo de 2008[68] sobre esos mismos restos clovis, aplicó modelos matemáticos para cuantificar qué suponía realmente ese 18% de restos con evidencias de caza activa de proboscídeos en comparación con otros hallazgos de caza de proboscídeos en yacimientos arqueológicos de todo el planeta. El análisis descubrió que el registro de los clovis mostraba que estos cazadores tenían en realidad, junto con los cazadores de la península ibérica, la frecuencia más alta registrada hasta la fecha de explotación prehistórica de proboscídeos como fuente de alimento de todo el mundo.[68]
Utilizando modelos cuantitativos y probabilísticos, basados en la profundidad temporal de los eventos de extinción, la demografía humana y el sesgo tafonómico, un estudio de 2012 de los antropólogos norteamericanos Surovell y Grund evaluaba cuántas asociaciones con fauna extinta deberían haber sido encontradas en la actualidad en Australia, Norteamérica y Nueva Zelanda. Concluían que debido al deterioro de los yacimientos a lo largo del tiempo, tal evidencia debía ser muy rara en Australia, de abundancia intermedia en Norteamérica y común en Nueva Zelanda, una conclusión que coincide de hecho con el estado actual del registro arqueológico.[62] Llegaron a una conclusión similar utilizando un análisis de la frecuencia relativa de las fechas de radiocarbono de cada región datadas en la época de coexistencia de los seres humanos y la fauna extinta. Argumentaban así que la escasez de evidencias de explotación de la fauna extinta no solo concordaba con la hipótesis de la caza excesiva, sino también con las demás hipótesis de extinción que se han propuesto, por lo que concluían que la crítica de la asociación es en realidad irrelevante para discriminar entre cualquiera de las distintas hipótesis.[62]
Primeros análisis estadísticos globales de las causas de la extinción (2012-2015)
Hasta la segunda década del SXXI, los estudios relativos a las extinciones cuaternarias se venían centrando en determinados continentes o habían utilizado dataciones de baja fiabilidad. La datación fósil permite el establecimiento de una sincronía entre los eventos de extinción y sus potenciales causas.[1] En los últimos años, se ha publicado y revisado un número creciente de dataciones en todo el mundo.[1] Se han desarrollado también modelos climáticos mejorados para los últimos 122.500 años.[69] Estos avances han hecho posible por primera vez el análisis cronológico global de los cambios climáticos, la llegada humana a cada región y la extinción de taxones megafaunísticos abriendo un camino prometedor para resolver el debate sobre la extinción del Cuaternario.[1][23][33][27]
El problema de las dataciones erróneas
Las dataciones paleontológicas y arqueológicas son muy sensibles a los errores metodológicos.[70] La contaminación de la muestra, la escasez de los materiales, las interpretaciones erróneas de la estratigrafía, los métodos inadecuados de datación y otros problemas, pueden poner seriamente en peligro la exactitud de una fecha.[1] Para identificar datos fiables, muchos autores han utilizado diferentes escalas cuantitativas basadas principalmente en el material de muestra, asociaciones estratigráficas y el tipo de equipo y logística utilizado en cada estudio dado.[71][72][60][73] Las fechas provenientes de estos estudios han sido incorporadas a otros estudios posteriores sin que se hayan hecho nuevas valoraciones sobre su fiabilidad. En la mayoría de los casos, sin embargo, estas fechas carecen de cualquier tipo de ponderación sobre su exactitud, haciendo del filtrado de datos una necesidad ineludible para obtener conclusiones fiables.[1]
Primeros análisis globales con dataciones de alta fiabilidad (2013 y 2015)

Así en 2013 se publica el primer análisis global realizado exclusivamente con dataciones de alta fiabilidad por Bernardo Araujo.[6][33] Este trabajo estadístico aplicaba a nivel global —considerando las diferentes regiones donde hubo extinciones masivas de megafauna asociadas— modelos matemáticos para comparar la probabilidad de influencia de las dos variables (llegada humana y cambio climático), y lo hacía por primera vez exclusivamente a partir de dataciones de alta fiabilidad.[1] Para las fechas basadas en radiocarbono, los datos fueron filtrados utilizando la escala Mead-Meltzer[71] modificada en 2010 por Barnosky y Lindsey,[60] y aplicando criterios muy estrictos: para aceptar fechas paleontológicas y arqueológicas, tenían que alcanzar respectivamente un rango de 11 (de un rango máximo de 12), y de 13 (de un rango máximo de 17).[1] Por otra parte, dada su antigüedad, la mayoría de las dataciones realizadas en Oceanía durante su período de extinción masiva de la megafauna, se basan en métodos diferentes, principalmente U / Th (datación Uranio-Torio), OSL (datación luminescente ópticamente estimulada) y ESR (barrido de resonancia electrónica). Puesto que no existen sistemas de puntuación capaces de evaluar la exactitud de las fechas obtenidas por estos métodos, se diseñaron escalas clasificadas a lo largo de la escala Mead-Meltzer para evaluar la fiabilidad de las fechas U / Th y OSL. La datación ESR implica un sistema más complejo de técnicas, que hace que sus fechas sean más difíciles de adaptar a un sistema de puntuación simple. Por lo tanto, solo se incluyeron las dataciones en las que se había utilizado el CSUS-ESR (Sistema cerrado de series-U ESR), una variante más exacta del método ESR.[74][75]

Las regiones incluidas en el análisis fueron Australia, Tasmania, Europa meridional, Europa septentrional, Rusia central, Japón, Siberia occidental, Beringia, Norteamérica occidental, Norteamérica central, Norteamérica oriental, la región andina (Andes y costa occidental de América del Sur), el norte de América del Sur, el centro de América del Sur, la Patagonia , el Caribe, la isla de Wrangel, Madagascar y Nueva Zelanda. Solo se utilizaron fechas calibradas, consideradas fiables de acuerdo con la escala modificada de Mead-Melzer.[60] La base de datos utilizada en el estudio incluía más de 2000 fechas megafaunísticas (para 67 géneros) y más de 700 fechas arqueológicas para el Homo sapiens.
El estudio ofreció como resultado que los fenómenos de extinción estaban más cerca de lo esperado por efecto del azar al momento de la llegada del ser humano en 76 casos (cada caso es la extinción de un género en una de las 19 regiones del planeta estudiadas). El momento de la extinción fue más cercano al esperado por efecto del azar a los periodos de intenso cambio climático en solo 2 casos (un valor cercano al esperado para una ordenación aleatoria de las extinciones a lo largo del tiempo geológico). Finalmente las extinciones estaban más cerca de lo esperado por efecto del azar tanto a la llegada del hombre como a épocas de intensa variación climática en 8 casos; y a ninguna de estas variables en 40 casos, de un total de 126 casos.
Un segundo estudio[1] publicado en 2015 por Araujo y un equipo formado por otros cuatro especialistas en ecología y paleobiología profundizaba aún más en la exigencia de alta fiabilidad de los datos analizados, siendo el más riguroso publicado hasta la fecha, utilizando solo restos fósiles de datación de alta resolución y los datos climáticos más precisos, obtenidos a partir de catas de hielo en Groenlandia y la Antártida. Después del filtrado de datos, se realizó una calibración de las dataciones. Como última precaución, se hicieron correcciones de arranque en las fechas paleontológicas de América del Sur (utilizando la Cueva del Milodón, en Chile, como referencia) para evitar posibles sesgos causados por el efecto Signor-Lipps, siguiendo la metodología establecida por Barnosky Y Lindsey.[60] El uso de datos corregidos no afectó significativamente a los resultados, por lo que los autores optaron por utilizar datos no corregidos para realizar todos los análisis descritos en la siguiente sección, teniendo en cuenta que esto podría hacer que sus análisis fueran más conservadores a la hora de encontrar una asociación entre la llegada humana y la extinción megafaunística.
La llegada del hombre como causa necesaria de la extinción

El equipo de Araujo examinó un total de 2088 fechas para 67 géneros de megafauna extinta (58 mamíferos, 8 aves y 1 reptil), totalizando 126 unidades de muestreo independientes en las 19 regiones citadas. De forma similar, 762 fechas humanas cumplieron con los requisitos de fiabilidad del estudio. Los modelos mostraron que la gran mayoría de las extinciones (85 de 126, o el 67.4%) ocurrieron alrededor del tiempo de llegada de los humanos a cada región, lo que encajaba con la estrecha brecha temporal entre las dataciones de alta resolución de aparición de megafauna y las dataciones de alta resolución de la primera aparición de los humanos en todo el mundo. Este patrón surgió incluso a pesar del uso de fechas no corregidas, que de hecho subestimaban el tiempo de coexistencia entre humanos y megafauna. No se produjo una correlación similar de fechas de extinción con tiempos de intenso cambio climático: solo el 17,5% (22 de 126) de las fechas de alta resolución de la última aparición de megafauna ocurrieron en períodos de intensa fluctuación de 18O o 2H. Entre estos, solo el 1,6% (2 de 126) de los géneros desaparecieron en períodos ligados supuestamente solo a una gran variación climática y no a la llegada humana. Por otra parte, 65 (51,6%) estaban más cerca de lo esperado por el azar a la llegada humana. Veinte casos (15,9%) se asociaron a ambos eventos. En los 39 casos restantes con los datos de alta fiabilidad disponibles en 2015, no apareció ninguna asociación estadísticamente significativa.[1]

Se generaron igualmente 41 modelos GLMM (modelo mixto lineal generalizado), que abarcaban una gama de intervalos de tiempo de entre 1000 y 5000 años. Cuando se utilizó cualquiera de los dos proxis de temperatura para representar la variación climática para todo el planeta, el proxy antártico de 2H detectó un efecto climático más intenso que el groenlandés de 18O, pero aun así la llegada humana tuvo un efecto aún más potente en ambos casos. Utilizando cada proxy para representar la variación climática en su propio hemisferio, el momento de mayor variación climática se relacionó significativamente con la extinción de la megafauna en solo 1 de los 41 intervalos. En marcado contraste, la fecha de llegada humana fue con diferencia el mejor predictor de la extinción de la megafauna, con un efecto significativo en 40 de los 41 intervalos, casi un 100%. En los modelos que consideraban la interacción entre ambos factores, el término de interacción fue significativo en solo 4 de los 41 contenedores. Por lo tanto, el GLMM proporcionó una evidencia significativamente sólida de los efectos antropogénicos frente a los climáticos, como causa de la extinción de megafauna del Cuaternario, y muy escasa evidencia de una acción sinérgica de estos dos factores.[1]
El estudio ofreció así una perspectiva cuantitativa ampliada sobre los papeles relativos de la variación del clima y los impactos humanos en el evento de extinción del Cuaternario tardío, con los impactos humanos como una determinante mucho más sólida en el número de géneros extintos que la variación climática. La extinción de cientos de especies en todo el planeta es un proceso tan complejo que los autores concluían que difícilmente podría esperarse que un solo factor explicara completamente cada aspecto de la misma. Era muy probable que factores causales como los impactos humanos y los cambios climáticos actuaran de diferentes maneras en diferentes lugares, para producir el resultado final actual, como ponían de manifiesto los resultados del equipo de Araujo. También concluían que dado que los análisis recientes eran por primera vez realmente fiables, se podía afirmar que ambos factores no habían tenido en absoluto igual importancia en el evento de extinción.[1]
Muchas extinciones megafaunísticos del Cuaternario Tardío ocurrieron por tanto en ausencia de cambios climáticos relevantes, pero rara vez, si es que hubo alguna, ocurrieron independientemente de la llegada del Homo sapiens. El estudio concluía así que las variaciones climáticas pudieron ser una causa contribuyente, que a veces podría haber ayudado a determinar dónde y qué especies se extinguieron, mientras que los impactos antropogénicos fueron la causa necesaria, sin la cual el evento de extinción del Cuaternario tardío nunca habría tenido lugar.[1]
La dispersión del ser humano por el planeta
Los humanos modernos se originaron en el África oriental y la mayor parte de su existencia como especie ha estado restringida a una parte relativamente pequeña del mundo.
África y Asia tropical

Las fechas más antiguas para el Homo sapiens son por lo menos de 160.000 años y las más antiguas fuera de África - en Oriente Medio - son de 40.000 años después, alrededor de 120.000 años atrás. Desde esta zona, los humanos modernos se dispersaron por el Asia Tropical, y permanecieron restringidos a estas tres regiones durante 70.000 años más, hasta hace aproximadamente 50.000 años.[76]
Oceanía
La primera nueva gran masa terrestre en ser colonizada fue el Sahul, en Oceanía. El Sahul comprendía Nueva Guinea, varias masas de tierra que hoy son islas pequeñas, Australia y Tasmania, unidas todas entre sí en aquella época por puentes de tierra. El Sahul fue alcanzado por los seres humanos hace aproximadamente 50.000 años. Puesto que los colonizadores venían de Asia, su desplazamiento sigue un patrón de norte a sur y de las costas hacia el centro, reflejado en el registro fósil: primero ocuparon la actual Nueva Guinea y tierras adyacentes, luego Australia y por último Tasmania hace 41.000 años. Por su parte Nueva Zelanda, aislada por grandes extensiones de océano, permanecería deshabitada hasta la llegada de los maoríes hace aproximadamente 7 siglos.[2][34][54][77]
El Paleártico
La expansión de los seres humanos modernos hacia las zonas templadas de Eurasia comenzó hace más de 40.000 años, pero fue un proceso mucho más lento y largo que la conquista del Asia tropical y Oceanía, ya que los seres humanos parecen haber llegado a las zonas frías del gran continente eurasiático, como el norte de Siberia, hace menos de 20.000 años. Era la primera vez que los humanos modernos tenían que colonizar un continente contra un gradiente de temperatura -de un clima muy cálido a uno extremadamente frío - y posiblemente este factor esté detrás de que el proceso de expansión fuera tan largo.[33]
Las Américas
.jpg.webp)
Una vez que los seres humanos modernos desarrollaron un acervo cultural y tecnológico que les permitía sobrevivir a los inviernos extremadamente fríos del Paleártico, alcanzaron el extremo este de Eurasia, desde donde tenían acceso libre a las Américas a través del puente de Beringia. América del Norte se alcanzó al menos 15.000 años atrás. En ese momento, la mitad septentrional de América del Norte todavía estaba cubierta por una gruesa capa de hielo, ya que la deglaciación no había terminado aún. En consonancia el avance a través de esta parte del continente parece haber sido lenta, pero la primera evidencia de que los seres humanos habían alcanzado las llanuras centrales de Norteamérica es de hace aproximadamente 13.500 años. En las Américas, y especialmente en América del Sur, la frontera de colonización parece haberse expandido a velocidades muy diferentes en diferentes direcciones. Las fechas arqueológicas más antiguas en Sudamérica son de Monte Verde, cerca de Puerto Montt, en el sur de Chile, de hace más de 14.000 años. Esta fecha parece coherente con la hipótesis de que la ruta de colonización más rápida fue a lo largo de la costa del Pacífico, que no presentaba obstáculos geográficos. Al este de la barrera biológica de los Andes, con una geografía más compleja y densos bosques, la expansión humana parece haber sido mucho más lenta que en la costa oeste.
Las islas
Los últimos lugares a los que llegaron los seres humanos fueron las islas de todo el mundo, ya que alcanzarlas solo se hizo posible tras el desarrollo de la navegación y la invención de embarcaciones eficientes, capaces de atravesar los mares. Las excepciones fueron actuales islas como Tasmania o el archipiélago de Japón, que eran penínsulas conectadas a los continentes en el Pleistoceno tardío y que fueron colonizadas a pie poco después de que los humanos llegaran a la masa continental principal. Las primeras verdaderas islas oceánicas que fueron alcanzadas por los seres humanos fueron las Antillas del Caribe en las Américas, hace tan solo unos 6.000 años. Las principales islas del Mediterráneo parecen haberse alcanzado poco después, y Wrangel, el hogar de los últimos mamuts, fue ocupada hace unos 3.700 años, coincidiendo con la extinción de la última población mundial estos animales.[49][79] Madagascar, a pesar de ser una de las islas más grandes del mundo, solo fue colonizada por los seres humanos hace 2.300 años. Finalmente, Nueva Zelanda fue la última gran masa terrestre del mundo en ser colonizada por los humanos, hace apenas 700 años.[1][77]
Correlación entre la llegada de los humanos y las extinciones masivas
La notable correlación entre estas fechas de llegada humana y las fechas de las extinciones en las diferentes masas terrestres en todo el mundo ha sido señalada por numerosos autores, siendo el primero en hacerlo el geólogo Paul S. Martin en 1967, cuando expuso su hipótesis basándose en que había una coincidencia temporal cada vez más evidente entre la expansión del Homo sapiens por el planeta y los eventos de extinción masiva que afectaban de forma casi exclusiva a la megafauna,[35][8][29] una extinción

de megafauna que además no había ocurrido nunca con anterioridad al evento de extinción del Cuaternario, ya que todas las extinciones masivas anteriores habían afectado también a infinidad de otros grupos de seres vivos y la estructura general de las comunidades de mamíferos había persistido durante más de medio millón de años antes de la llegada del ser humano moderno.[13]
Desde entonces esta correlación ha sido señalada por numerosos científicos como evidencia de que el hombre moderno fue el principal causante de la extinción.[1][2][6][7][9][10][11][12][13][14][15][16][17][18][19][20][21][22][36][80] Es a partir de 2013 cuando el trabajo estadístico de Araujo[33] inicia una nueva etapa en la que se desarrollan metodologías de análisis comparativos que permiten calcular la probabilidad de cada causa de extinción mediante modelos matemáticos y dataciones de alta precisión. El estudio de Araujo y otros posteriores de la misma naturaleza están estableciendo una metodología[13][81][82] que viene poniendo de manifiesto en los últimos años cómo la llegada del hombre es la causa más probable de extinción de la megafauna a nivel global y/o regional, con estudios específicos que así lo indican para Oceanía[25] Asia[12] Madagascar[40] y América del Norte[13] entre otras regiones, y en ocasiones con un posible efecto no sinérgico sino contributivo y secundario[1] de cambios climáticos/florísticos y erupciones volcánicas que pudieron acelerar el proceso desencadenado por nuestra especie, como en el caso de Sudamérica.[61][60] Si bien el cambio climático fue una causa contribuyente que ayudaría a explicar el proceso de extinción de ciertas especies, los impactos antropogénicos serían la causa necesaria que lo impulsó.[1]
Desarrollo y alcance de las extinciones de megafauna del Cuaternario tardío
En este evento de extinción masiva se han perdido 2/3 de todos los géneros de mamíferos y la mitad de la megafauna terrestre (con un peso mayor de 44 kilos) de la Tierra.[6][7]
Asia tropical

Aunque tradicionalmente se ha incluido el Asia tropical como la segunda región junto al África subsahariana donde no ha habido extinciones masivas de megafauna, lo cierto es que esta zona ha sufrido pérdidas megafaunísticos muy notables que se han pasado por alto en muchos estudios porque no muestran un pico claro en el Cuaternario Tardío y porque algunas especies emblemáticas como el elefante asiático, el tigre o el rinoceronte indio han sobrevivido hasta nuestros días.[84][37]
Sin embargo el registro fósil muestra que toda la región, incluidas todas las islas grandes desde Indonesia a Nueva Guinea, y muchas más

islas pequeñas, albergaron más especies grandes de vertebrados y mucha más biomasa de megavertebrados en el Pleistoceno de las que albergan en la actualidad[86][87] con muchas especies viendo su rango de distribución original reducido a una mínima fracción a lo largo de los últimos 50.000 años —al 10% de su rango máximo de distribución en el Holoceno y aún más desde el Pleistoceno.[84] Además varias especies de megafauna se han extinguido durante el evento de extinción del Cuaternario tardío, varias de ellas como el elefante chino o el tapir gigante, ya en el Holoceno:
- Elefantes:
- Elefante de Naumann (Palaeoloxodon naumanni)[39][88][89]
- Elefante chino (Stegodon orientalis)[90][91]
- Elefante de Indonesia (Stegodon trigonocephalus)[90]
- Rinocerontes:
- Pangolín gigante asiático (Manis paleojavanica)[84]
- Uro asiático (Bos primigenius primigenius)
- Equus sp.[90]
- Jabalí (Sus lydekkeri)[90]
- Orangután (Pongo sp.)[90]
- Tapir gigante (Megatapirus augustus)[90]
- Hiena manchada asiática (Crocuta crocuta ultima)[90][92]
- Hiena gigante (Pachycrocuta brevirostris)[90][93]
- Tortuga gigante de Timor (Megalochelys sp.)[38]
El Asia tropical soporta un episodio de extinción masiva en curso de megafauna que amenaza a las especies supervivientes.[94]
Japón

Durante los períodos glaciales, las actuales islas de Sajalín y Hokkaido estaban conectadas al continente asiático formando una península. Sin embargo, el estrecho de Tsugaru que separa Hokkaido y Honshu tiene unos 130 m de profundidad, similar a la profundidad del estrecho de Tsushima que separa la península coreana y el sur de la entonces península japonesa. Así, solo durante los máximos glaciales existiría una conexión terrestre entre las actuales islas de Hokkaido y Honshu. La mayor parte de la megafauna que desapareció durante el Pleistoceno Tardío y casi todas las pruebas actuales de dispersiones tempranas de homínidos en Japón se limitan a esa región meridional.[39] Japón fue colonizado por los humanos desde fechas tan tempranas como unos 50.000 años atrás,[39] aunque la fecha de 35.000 años es generalmente la más aceptada.[95] La megafauna que aparece con mayor frecuencia en los yacimientos estudiados son los mamutes, el elefante de Naumann y los ciervos gigantes asiáticos del género Sinomegaceros. A partir de las evidencias de caza directa y/o descuartizamiento encontrados en los fósiles de estos tres taxones, un estudio de 2009 dedujo que el proceso de extinción pasó por tres etapas en Japón:
• El proceso de extinción de Palaeoloxodon, Sinomegaceros y Mammuthus en Japón se inició ya tras la primera colonización humana del archipiélago japonés, hace menos de 50.000 años, como evidencian las interacciones humano-megafaunísticos halladas en yacimientos como Tategahana. Sin embargo, las dispersiones humanas iniciales en el archipiélago fueron probablemente de pequeña escala, como indican la baja densidad de yacimientos arqueológicos y la baja densidad de evidencias de interacción en cada uno de los yacimientos supuestamente fechados entre 50.000 y 30.000 años atrás.[39]
• Después del límite de 30.000 años, más de 5.400 yacimientos arqueológicos paleolíticos aparecen en Japón. Esto refleja directamente el aumento significativo de la densidad de población humana y la proporción de las islas utilizadas, lo que habría influido directa o indirectamente en los rangos de distribución locales de la megafauna. En estos yacimientos aparecen huesos de proboscídeos con marcas de corte y restos de puntas de cuarzo incrustados en ellos.[39]
• Muchas de las extinciones probablemente ocurrieron después de los 30.000 años y durante la transición MIS 3-2 (entre 30.000 y 20.000 años atrás), en lugar de durante la transición MIS 2-1 (entre 15.000 y 10.000 años atrás) y antes del advenimiento del período Jomon . De hecho, actualmente no hay yacimientos conocidos de Jomon que hayan dado evidencia de restos óseos de Palaeoloxodon, Mammuthus o Sinomegaceros.[39]
Sahul (Australia y Nueva Guinea)

Todos los mamíferos terrestres, reptiles y aves australianos que pesaban más de 100 kilogramos, y seis de los siete géneros con una masa corporal de 45 a 100 kilogramos, desaparecieron.[83] En total 23 de 24 géneros de megafauna devinieron extintos.[83] La determinación, tanto del momento de extinción de la megafauna como de la primera llegada humana a Sahul, ha sido polémica debido a la falta de datos fiables y sesgos introducidos por el efecto Signor-Lipps. Teniendo en cuenta estas deficiencias, los resultados de un estudio llevado a cabo en 2015 por 18 autores de diversas universidades con dataciones de alta fiabilidad[25] respaldaban las conclusiones generales de estudios previos[83][96] Los resultados fueron mucho más precisos gracias al uso de conjuntos de datos más diversos y ampliados que fueron rigurosamente analizados utilizando las herramientas analíticas más eficientes hasta la fecha.[25]

Se acepta generalmente que los seres humanos estaban presentes en la actual Australia hace unos 48.000 años.[97][98] La estimación de edad modificada de 55.600 años que obtenía el estudio de 2015 reflejaba la temprana colonización humana del norte de Australia,[96][99] pero los vacíos geográficos en datos de alta calidad impiden una interpretación fiable del patrón de dispersión humana a través de Sahul. Está claro, sin embargo, que los seres humanos estaban presentes en Tasmania hace unos 39.000 años[97] y en el centro árido de Australia unos 35.000 años atrás.[100]
La ventana de 13.500 años de coexistencia entre humanos y megafauna estimada es suficientemente amplia para que grupos de cazadores-recolectores se dispersaran y se establecieran a través de Sahul[101] extinguiendo muchas especies de megafauna sin una ola de extinción rápida y continental. Estos resultados no descartaron la posibilidad de una extinción masiva rápida por los humanos a escalas más locales o regionales.[102]
Se ha estimado que la caza de baja intensidad de especies con una tasa de reproducción baja (como la caza de un solo juvenil por persona por década) pudo resultar en que una especie se extinguiera después de unos pocos cientos de años de aprovechamiento.[34] Esta caza selectiva y de baja intensidad sería virtualmente imperceptible en el registro arqueológico, lo que junto a la pérdida tafonómica a lo largo del tiempo explica la ausencia de yacimientos de despiece. A pesar de ello en noviembre de 2016, se publica el primer hallazgo con pruebas de cacería humana de megafauna en Australia.[65] En el registro fósil de cabo Pasley, el colapso de la población megafaunística comenzó 2.000 años después de la fecha establecida para la dispersión humana en Australia en unos 47.000 años.[102]



Los modelos cuantitativos demostraban que incluso grupos pequeños de cazadores-recolectores que viven en un vasto continente y que utilizan tecnologías basadas en la piedra podían exterminar exitosamente especies con bajas tasas de reproducción, como los mamíferos de gran tamaño. Esta conclusión no podría ser apoyada por resultados basados solo en una lectura directa del registro fósil, ya que sería sesgada por el efecto Signor-Lipps no corregido.[103] Al incluir solo restos de megafauna y registros arqueológicos fiablemente datados en los modelos del estudio —y corrigiendo explícitamente el efecto Signor-Lipps— la duración de la coexistencia entre la megafauna humana y el ser humano pudo estimarse con un intervalo de confianza estadística mucho mayor que en estudios anteriores. El enfoque del estudio permitió así discriminar entre los diversos mecanismos de extinción propuestos para la última megafauna cuaternaria de Sahul y reveló que el cambio climático no fue un causante de la extinción continental de estos géneros.[25][104]
Un segundo estudio con dataciones de alta precisión venía a reforzar sus resultados en 2017.[102]
Entre la megafauna perdida en el evento de extinción están los siguientes taxones:
- Todos los miembros de la familia Diprotodontidae:
- Diprotodon optatum (ocupaban un nicho ecológico similar al de los proboscídeos en el resto del planeta, parientes gigantes de los wombats)[104]
- Diprotodon minor[105]
- Hulitherium tomasetti (un gran marsupial herbívoro)[84]
- Maokopia ronaldi (diprotodonto)[83]
- Rinoceronte marsupial (Zygomaturus trilobus)[83]
- Varios canguros gigantes de la familia Macropodidae:
- Ualabí gigante (Wallabia kitcheneri)
- Canguros gigantes de patas largas Macropus:
- Canguros gigantes de hocico corto del género Procoptodon:
- Procoptodon goliath[83]
- Procoptodon sp. indet.[83]
.jpg.webp)

- Canguro omnívoro (Propleopus oscillans)[83]
- Ualabís gigantes del género Protemnodon:[104]
- Canguros gigantes del género Simosthenurus:[104]
- Canguros gigantes del género Sthenurus:
- Uombat gigante (Phascolonus gigas)[83]
- Phascolomys major[105]
- Phascolomys medius[105]
- Uombat (Vombatus hacketti)[83]
- Tapir marsupial (Palorchestes azcal)[83]
- Koala gigante (Phascolarctos stirtoni)[105]
- Rata canguro gigante (Borungaboodie hatcheri)[83]

- Equidna gigante (Megalibgwilia ramsayi )[83]
- Equidna gigante (Zaglossus hacketti)
- León marsupial Thylacoleo carnifex (un carnívoro del tamaño de los mayores felinos actuales)[104]
- Tigre de Tasmania (Thylacinus cynocephalus, extinto primero en Nueva Guinea, luego en Australia, y finalmente en Tasmania en el SXX)[83]
- Diablos de Tasmania gigantes:
- Ave gigante galliforme (Genyornis newtoni)[104]
- Ave gigante (Progura naracoortensis)[83]
- Varano gigante (Varanus priscus o Megalania prisca)[104]
- Todas las tortugas terrestres gigantes del género Meiolania sp.[83]
- Tortuga terrestre gigante (Ninjemys oweni)[83]
- Todos los cocodrilos del género Paludirex sp.[83]
- Cocodrilos terrestres del género Quinkana sp.[83]
- Serpiente constrictora (Wonambi naracoortensis)[83]
Europa y el Asia Paleártica

A lo largo del Pleistoceno medio y superior la fauna europea experimentó repetidas expansiones y contracciones alternas en sus rangos de distribución, con las fases frías favoreciendo un conjunto de especies y las templadas el otro. En cada etapa fría los animales interglaciales retrocedieron al sur de Europa, y recolonizaron Europa central y septentrional en las etapas cálidas.[106] Por el contrario, especies extendidas en las fases frías, se retiraron hacia el norte y/o hacia la estepa, o a altitudes más altas durante los interglaciales. Este ciclo se interrumpió repentinamente en los últimos 50.000 años aproximadamente, coincidiendo con un incremento sin precedentes de las extinciones megafaunísticas.[106]
En consonancia con el lento avance del Homo sapiens en el Paleártico, las extinciones paleárticas de megafauna aparecen más dispersas en el tiempo, presentando tres pulsos principales, uno en la Eurasia holártica meridional, el segundo en la Eurasia holártica septentrional, y el tercero y último en las islas mediterráneas y siberianas.[107] En la mitad sur, la zona más próxima a la cuenca mediterránea, varias especies de megafauna como el elefante europeo o el hipopótamo común desaparecieron de tierras continentales hace entre 45.000 y 20.000 años, sobreviviendo remanentes de elefantes, hipopótamos y otras especies de megafauna en las islas mediterráneas, que no serían alcanzadas por el hombre hasta mucho después, con el desarrollo de la navegación.[53][6] Las extinciones en masa en latitudes septentrionales constituyeron el segundo pulso y llegaron miles de años después, de 14.000 a 9.000 años atrás, de nuevo con poblaciones remanentes de mamut lanudo en la Isla de Wrangel que sobrevivieron hasta hace 3.800 años, coincidiendo con los primeros restos humanos.[49]
La colonización de Eurasia fue el único momento en la dispersión del Homo sapiens en el que los humanos paleolíticos, hasta entonces una especie completamente tropical, se vieron obligados a avanzar frente a ambientes cada vez más fríos a medida que expandían sus poblaciones hacia latitudes más altas. Las fechas de primera aparición del ser humano en Rusia Central y Europa tienen más de 45 mil años de antigüedad, unos 30.000 años más antiguas que las de Bering y casi 20.000 años más que las primeras fechas humanas en el Ártico.[108] En consecuencia, Surovell en 2009,[26] después de corregir el sesgo tafonómico, encontró evidencias de que los mamuts sufrieron un largo proceso de progresivo enrarecimiento hasta la extinción, correlacionado con el aumento de las densidades humanas.[26] Así, el largo proceso de poblamiento de Eurasia septentrional contra el gradiente de frío puede haber permitido una mayor brecha temporal entre las primeras fechas de aparición de los humanos modernos y las últimas fechas de aparición de la megafauna. Otro factor que probablemente influyó en que las extinciones fueran más lentas en Eurasia es la coexistencia de la megafauna con especies de Homo anteriores durante varios miles de años antes de la llegada de los humanos modernos.[1][109] Homo heidelbergensis y Homo neanderthalensis habían estado presentes allí durante cientos de miles de años antes de la diáspora de los humanos modernos a Eurasia y ya cazaban megafauna.[110] Así, similar a lo ocurrido en África, la coevolución posiblemente concedió a los animales grandes una cierta resistencia a la caza por humanos, aunque insuficiente para evitar la extinción de 21 de sus 37 géneros de megafauna tras la llegada del Homo sapiens,:

- Mamut lanudo (Mammuthus primigenius)
- Elefante europeo (Palaeoloxodon antiquus)
- Rinoceronte lanudo (Coelodonta antiquitatis)
- Rinoceronte de Merk (Stephanorhinus kirchbergensis).
- Rinoceronte de hocico estrecho (Stephanorhinus hemiotoechus)
- Rinoceronte elasmoterio (Elasmotherium sibiricum)
- Hipopótamo africano (Hippopotamus amphibius).[111]
- Hipopótamo europeo (Hippopotamus antiquus)
- Ciervo gigante (Megaloceros giganteus)
- Ciervo gigante (Eucladoceros)
 Cráneo fósil del ciervo gigante Eucladoceros, género de ciervos gigantes europeos extintos en el evento de extinción del Cuaternario.
Cráneo fósil del ciervo gigante Eucladoceros, género de ciervos gigantes europeos extintos en el evento de extinción del Cuaternario. - Alce de frente ancha (Cervalces latifrons)
- Bisonte de estepa (Bison priscus)[112]
- Bisonte europeo (Bison bonasus, extinto en estado salvaje en el SXX)[113]
- Toro etrusco Leptobos sp.
- Uro (Bos primigenius primigenius)[114]
- Buey almizclero (Ovibos moschatus, extinto en Eurasia)
- Tarpán (Equus ferus ferus)
- Asno europeo (E. hydruntinus)
 Elasmotherium , rinoceronte de un solo cuerno extinto en Eurasia en el evento de extinción del Cuaternario.
Elasmotherium , rinoceronte de un solo cuerno extinto en Eurasia en el evento de extinción del Cuaternario. - Antílope de cuernos de tornillo Spirocerus kiakhtensis[115]
- Gacela persa Gacela subgutturosa
- Camello europeo, (Camelus knoblochi)[116]
- Nutria robusta europea (Cyrnaonyx)
- Nutria mediterránea (Algarolutra)
- Leopardo europeo (Panthera pardus spelaea)
- Cuón alpino europeo (Cuon alpinus europaeus)
- Gato cimitarra (Homotherium sp.)
 Homotherium desapareció de Eurasia y Norteamérica a finales del Pleistoceno, en el evento de extinción del Cuaternario.
Homotherium desapareció de Eurasia y Norteamérica a finales del Pleistoceno, en el evento de extinción del Cuaternario. - Lynx issiodorensis
- Lince mediterráneo (Lynx spelaeus)[117]
- León de las cavernas (Panthera leo spelaea)
- Hiena de las cavernas (Crocuta crocuta spelaea)
- Osos Ursus:
- Oso de Gamssulzen (Ursus ingressus)[118]
- Oso de las cavernas menor (Ursus rossicus)
- Oso de las cavernas (Ursus spelaeus)
- Oso polar gigante (Ursus maritimus tyrannus)
- Avestruz asiático (Struthio asiaticus)[119]
América del Norte

En América del Norte, el proceso de extinción fue comparativamente "rápido", las poblaciones megafaunísticas se derrumbaron a lo largo de casi 1000 años, de 14.800 a 13.700 años atrás, durante el período cálido de BøllingAllerød.[56] La mayoría de las extinciones finales tuvieron lugar entre 13.000 y 11.500 años atrás,.[120] De nuevo, algunos géneros sobrevivieron hasta tiempos más recientes, como los mastodontes, cuyos últimos registros, en la región de los Grandes Lagos, datan de hace 9.900 años.[121]
La megafauna norteamericana antes de la extinción era extremadamente rica y bastante similar a la fauna actual de África.[122] Entre otros, había mamuts lanudos, mamut colombinos, mastodontes, perezosos gigantes, camellos americanos, caballos y bisontes, incluyendo el enorme bisonte de cuernos largos. Además, América del Norte fue el hogar de una fauna carnívora excepcionalmente rica, incluyendo el oso gigante, el tigre dientes de sable, el gato cimitarra, dos leones (Panthera atrox y Panthera spealea), lobos gigantes y guepardos americanos. Las extinciones en las Américas supusieron la eliminación de todas las especies de mamíferos por encima de los 1000 kg, incluyendo los que habían emigrado desde América del Sur en el Gran Intercambio americano. En América del Norte, la aparición del pulso de extinción principal y la primera aparición de los paleoamericanos Clovis son virtualmente indistinguibles en el tiempo.[6][11][13][20]
Los anteriores pulsos de extinción ocurridos en Norteamérica no fueron comparables al de la extinción cuaternaria, porque no afectaron de forma casi exclusiva a la megafauna, e implicaron principalmente reemplazos de especies dentro de nichos ecológicos, mientras que el último resultó en que muchos nichos ecológicos quedaran por primera vez vacíos. Los animales que aparecen con mayor frecuencia en los puntos de descuartizamiento de los Clovis son los proboscídeos, de donde se deduce que mamutes y mastodontes fueron su principal presa.[17] Un estudio de 2008 sobre los restos Clovis aplicó modelos matemáticos para elaborar un análisis comparativo y poder así cuantificar qué suponía que el 18% de los restos conocidos de esta cultura presentaran evidencias de caza activa en comparación con otros hallazgos en yacimientos de todo el planeta. El análisis descubrió que el registro de los Clovis mostraba de hecho que estos cazadores tenían la frecuencia más alta registrada hasta la fecha de explotación prehistórica de proboscídeos como fuente de alimento de todo el mundo, junto con los cazadores de la península ibérica.[17]
Las extinciones de América del Norte incluyeron:





_1_(15257877377).jpg.webp)


- Todos los elefantes de la familia Gomphotheriidae:
- Todos los mamuts:
- Mamut lanudo (Mammuthus primigenius)
- Mamut colombino (Mammuthus columbi)
- Mamut pigmeo (Mammuthus exilis)
- Mastodonte americano (Mammut americanum)
- Todos los équidos:
- Equus alaskae
- Caballo mexicano (Equus conversidens)
- Equus francisci
- Tarpán (Equus ferus ferus)
- Equus fraternus
- Equus giganteus
- Asno salvaje del Yukón (Equus lambei)
- Caballo occidental (Equus occidentalis)
- Equus semiplicatus
- Caballo de Hagerman (Equus simplicidens)
- Caballo de Scott (Equus scotti)
- Tapir de California (Tapirus californicus)
- Tapir de Merriam (Tapirus merriami)
- Tapirus veroensis
- Camello occidental (Camelops hesternus)
- Llama norteamericana (Hemiauchenia)
- Llama de patas gruesas (Paleolama)
- Antílope Capromeryx
- Antílope Stockoceros
- Antílope Tetrameryx
- Ciervo de montaña americano (Odocoileus lucasi)
- Ciervo-alce (Cervalces scotti)
- Buey de los arbustos (Euceratherium collinum)
- Búfalo americano (Bootherium bombifrons)
- Cabra de montaña de Harrington (Oreamnos harringtoni)
- Bisonte gigante (Bison latifrons)
- Bisonte occidental (Bison occidentalis)
- Castores gigantes (Castoroides ohioensis y Castoroides leiseyorum)
- Saiga (Saiga tatarica, extinto en Norteamérica)
- Tortugas gigantes (Hesperotestudo sp. y Gopherus donlaloi)
- Todos los buitres gigantes teratornítidos (Teratornithidae):
- Aiolornis incredibilis
- Cathartornis gracilis
- Oscaravis olsoni
- Teratornis merriami
- Teratornis woodburnensis
- Gymnogyps amplus
- Buitres del Nuevo Mundo del género Breagyps sp.
- Buitres del Nuevo Mundo del género Neogyps sp.
- Vaca marina de Steller (Hydrodamalis gigas)
- Neochoerus (como el capibara de Pickney, N. pinckneyi)
- Hutias gigantes (Heptaxodontidae)
- Pika gigante (Ochotona whartoni)
- Todos los perezosos gigantes:
- Eremotherium,
- Glossotherium
- Megalonyx
- Neocnus
- Nothrotheriops
- Nothrotherium
- Paramylodon
- Todos los armadillos gigantes:
- Glyptotherium
- Doedicurus
- Armadillo bello (Dasypus bellus)
- Pampathéridos (como el género Holmesina)
- Mofeta de hocico corto (Brachyprotoma obtusata)
- Osos de cara corta (Arctodus simus y Arctodus pristinus, unos de los mayores mamíferos carnívoros terrestres de todos los tiempos)
- Oso de las cavernas de Florida (Tremarctos floridanus)
- Gato cimatarra (Homotherium serum)
- Tigre de dientes de sable (Smilodon fatalis)
- Guepardo americano (Miracinonyx)
- León americano (Panthera atrox)
- León de las cavernas (Panthera leo spelea)
- Tigre siberiano (Panthera tigris altaica)
- Jaguar norteamericano del Pleistoceno (Panthera onca augusta distribución semirecolonizada por otra subespecie)
- Margay (Leopardus weidii, extinto localmente)
- Ocelote (Leopardus pardalis, extinto localmente, área de distribución recolonizada marginalmente)
- Cuón (Cuon alpinus)
- Lobo gigante (Canis dirus)
- Coyote del Pleistoceno (Canis latrans orcutti)
- Pecarí de cabeza plana (Platygonus)
- Pecarí de nariz larga (Mylohyus)
- Pecarí de collar (Pecari tajacu, extinto localmente, área reconolizada parcialmente)
América del Sur








Tomando las fechas de radio-carbono en su valor nominal y asumiendo que el desfase entre la última desaparición datada y la desaparición final real es similar a través de taxones y continentes, el pulso de extinción comenzó en América del Sur unos 1000 años después de que los humanos llegaran por primera vez al continente.[6] En América del Sur, las extinciones se extendieron a lo largo de un período de tiempo más largo que en América del Norte, desde aproximadamente 13.000 años atrás hasta hace 7.800 años.[60] Esto supone que, aunque hay un considerable solapamiento de tiempos entre los continentes, muchas de las extinciones sudamericanas tuvieron lugar después de que el proceso se hubiera completado en su mayoría en Norteamérica.[6]
La megafauna sudamericana incluía al menos dos especies de elefantes de los géneros Haplomastodon y Cuvieronius, tigres dientes de sable, y una rica diversidad de perezosos de las familias Megatheridae, Megalonychidae, Milodontidae y Nothotheridae. Los perezosos más grandes de los géneros Megatherium y Eremotherium eran animales gigantescos y lentos que podían pesar más de cinco toneladas. El tamaño medio de la megafauna sudamericana era más grande que el de la megafauna africana.[58] Había grandes herbívoros nativos similares a los hipopótamos llamados toxodontes (Toxodon platensis), muchos gliptodontos gigantescos parecidos a los armadillos, y una rica diversidad de herbívoros con pezuñas, incluyendo caballos y animales tan peculiares como Macrauchenia. Aunque hoy en día América del Sur es relativamente pobre en megafauna —los tapires, los camélidos, los jaguares y los ciervos de pantanal son los pocos mamíferos grandes que quedan— esto no era así ante de la extinción, cuando contaba con una de las megafaunas más variadas y grandes del planeta.[60]
Los taxones extintos en el Cuaternario tardío fueron:
- Todos los elefantes sudamericanos:
- Elefantes del género Cuvieronius
- Elefantes del género Stegomastodon (=Haplomastodon)[125]
- Dientes de sable Smilodon fatalis
- Dientes de sable Smilodon populator
- Jaguar sudamericano del Pleistoceno Panthera onca mesembrina
- Jaguar Panthera onca augusta
- Osos sudamericanos Arctotherium
- Vampiro gigante Desmodus draculae
- Lobo gigante Canis dirus
- Cánido argentino Theriodictis platensis
- Cánidos Protocyon
- Zorros sudamericanos Dusicyon avus y Dusicyon cultridens
- Zorro vinagre del Pleistoceno Speothos pacivorus
- Armadillo Pachyarmatherium
- Armadillo Dasypus bellus
- Ciervo Antifer
- Agalmaceros[126][127]
- Camelidae
- Perezosos gigantes:
- Mylodontidae
- Catonyx
- Glossotherium
- Lestodon
- Mylodon
- Nematherium
- Octomylodon
- Orophodon
- Scelidotherium
- Megatheriidae
- Nothrotheriidae
- Nothropus
- Nothrotherium
- Megalonychidae
- Proplatyarthrus
- Valgipes gervais et Valgipes bucklandi[128]
- Mylodontidae
- Armadillos gigantes Glyptodontidae
- Doedicurus
- Eleutherocercus
- Glyptodon
- Heteroglyptodon[129]
- Hoplophorus
- Lomaphorus
- Neosclerocalyptus
- Neuryurus[130]
- Panochthus
- Parapanochthus[131]
- Plaxhaplous
- Sclerocalyptus
- Armadillos gigantes Pampatheres (e.g. Holmesina, Pampetherium et Tonnicinctus)[132]
- Armadillos gigantes Eutatus
- Propaopus[133]
- Neochoerus
- Equidae
- Equus (Amerhippus)
- Hippidion
- Onohippidium
- Toxodón Toxodon
- Mixotoxodon
- Liptopterna
- Mesotheriidae
- Caiman venezuelensis
Las Antillas

Las Antillas tenían antes de su extinción masiva, una megafauna muy diversa y similar a la continental en las Américas, y constituyen —junto a las islas mediterráneas, la Isla de Wrangel, la de Saint Paul, y Madagascar—, uno de los ejemplos más evidentes de cómo las extinciones masivas de megafauna ocurrieron miles de años más tarde en las islas que en los continentes, aun cuando a menudo albergaban las mismas especies, géneros o familias que se habían extinguido en tierra firme.
En el caso de las Antillas, el grupo más estudiado hasta la fecha es el de los perezosos gigantes, que se extinguieron miles de años después que los perezosos continentales. Cuando se examinan los depósitos más jóvenes de estiércol fósil de perezoso gigante, los de América del Sur son solo dos siglos más antiguos que los más recientes de América del Norte.[28] Los restos de perezosos gigantes están ausentes de los abundantes depósitos de fósiles del Holoceno en ambos continentes. En Cuba y Haití, por el contrario, 8 de las 13 fechas de C14 son del Holoceno en lugar del Pleistoceno, con dos de ellos de hace solo 5.000 años.[28]
Los yacimientos arqueológicos más tempranos de estas islas no han sido estudiados todavía intensivamente, pero varios emplazamientos han producido artefactos líticos (especialmente grandes hojas de chert) asociados con dataciones de C14 tan antiguas como 5.500 años.[28]
Las islas mejor estudiadas hasta la fecha son La Española y Cuba, donde se han descrito varias especies de megafauna destacando sus perezosos gigantes y varias rapaces gigantes.
Entre los taxones extintos se encuentran los siguientes:
- Todos los perezosos gigantes:
- Búho gigante cubano (Ornimegalonyx)
- Cóndor cubano (Gymnogyps varonai)
- Caracara sp.
Islas mediterráneas

La cuenca mediterránea incluye unas 370 islas (exceptuando islotes menores de 0,1 km²). Cerca de 300 de ellas son pequeñas islas (entre 0,1 y 100 km²). Junto con las 36 islas medianas (100 a 1000 km²), estas islas de tamaño discreto están situadas en su mayor parte cerca del continente o de las grandes costas insulares, a cuyas masas terrestres estaban unidas hasta el final de la última transgresión marina. Por otra parte, pocas de ellas han dado suficientes datos paleontológicos para una reconstrucción fiable de la historia de las relaciones hombre-animal.[53] Por estas razones, la mayoría de los estudios respecto a la megafauna y su extinción se han centrado en las grandes islas (más de 1000 km²), y especialmente en las que han permanecido completamente separadas del continente al menos desde el comienzo del Pleistoceno Superior. Durante los últimos 130.000 años, estas islas verdaderas no pudieron ser alcanzadas por especies de vertebrados no voladores o acuáticos. Solo hay seis islas que cumplan con estas características, repartidas entre las áreas oriental y occidental de la cuenca: Chipre, Rodas, Creta, Cerdeña, Córcega y Mallorca, aunque los datos de Rodas y Creta son aún demasiado escasos para extraer conclusiones respecto a las causas de su extinción.[53] Habiendo estado aisladas durante mucho tiempo en el Pleistoceno, todas estas islas fueron habitadas durante el último periodo glaciar por una megafauna muy diversa y extremadamente endémica, generalmente compuesta por ciervos, hipopótamos y elefantes,[53][134] y algunos roedores y búhos gigantes.[53][135]
.jpg.webp)
Con los conocimientos actuales, se considera que Homo sapiens no colonizó las grandes islas mediterráneas verdaderas antes del noveno milenio.[53] Se sabe que los elefantes habitaban hasta el Holoceno en numerosoas islas del Mediterráneo gracias a su capacidad para nadar grandes distancias utilizando la trompa a modo de esnórquel. Antes de que los humanos desarrollaran la navegación y alcanzaran las islas mediterráneas en el Holoceno, había varias especies de elefantes, mamutes e hipopótamos repartidos por la mayoría de las islas mediterráneas: Sicilia,[136] Malta,[137] Chipre,[138] Creta,[139] Tilos,[140] Cos,[141] Imbros,[142] Serifos,[143] Citnos,[144] Delos,[145] Naxos,[145] Citera,[144] y Rodas.[146] El último elefante enano, Elephas tiliensis o Palaeoxodon tiliensis, podría haber sobrevivido hasta tiempos históricos en Tilos, hasta hace 4000 años según las dataciones e incluso hasta unos 3500 años atrás según han apuntado algunos autores a partir de una pintura en la tumba del visir de Tutmosis III y Amenhotep II (del periodo de 1470 a 1445 AC).[52][147]
• Cerdeña

El Holoceno de Cerdeña está entre los peor documentados de las islas mediterráneas, debido a la escasa cantidad de restos faunísticos en las excavaciones arqueológicas, aunque a veces estos yacimientos sean numerosos, especialmente para la Edad de Bronce nurágica y la Antigüedad Clásica. El periodo glacial tardío, el Holoceno temprano y el Neolítico temprano son respectivamente conocidos únicamente por un yacimiento. Sin embargo la cueva de Corbeddu ha dado la secuencia estratigráfica y las asociaciones óseas más importantes para el periodo glacial tardío y el Holoceno temprano en las islas del Mediterráneo.[148][149][150] Una verdadera colonización del Pleistoceno superior seguramente habría producido una rápida extinción del ciervo gigante sardo Megaloceros cazioti, pero este todavía es abundante en el estrato correspondiente al noveno milenio de la cueva de Corbeddu.[53] Para Jean-Dennis Vigne, la cueva de Corbeddu debe ser considerada como un indicador fiable de la ausencia de seres humanos en Cerdeña durante el Paleolítico Tardío, de su aparición durante los milenios 9º-8º ya en el Holoceno, y probablemente de la persistencia de la megafauna endémica hasta ese momento, considerándose la causa probable de su extinción la llegada del hombre.[53] Cynotherium sardous y Megaloceros cazioti todavía están presentes en los niveles del 9º-8º milenio en la Cueva de Corbeddu, y por lo tanto asociados con los primeros huesos humanos de Cerdeña.[148][149][150] Sin embargo, ya están completamente ausentes del refugio mesolítico de Porto Leccio, del octavo milenio.[53] El gamo Dama dama ha sido sugerido como elemento megafaunístico para la Edad del Bronce.[151] La especie está atestiguada con seguridad durante la Baja Edad Media[152] y se extinguió en 1968.[151]
• Córcega
Con cerca de 100 conjuntos faunísticos y cerca de 200.000 huesos de mamíferos identificados, Córcega es probablemente la isla mediterránea mejor documentada para el Holoceno. Megaloceros cazioti y Cynotherium sardus son los únicos dos grandes mamíferos conocidos del Pleistoceno Superior de Córcega.[53] Los datos estratigráficos de la cueva de Grltulu han demostrado que este ciervo y cánido endémicos del archipiélago sardo-corso, sobrevivieron en Córcega hasta el final del glacial tardío, posiblemente hasta el comienzo del Holoceno, pero ya están completamente ausentes de las capas culturales del octavo milenio.[53] Numerosos datos paleontológicos, biológicos y arqueozoológicos indican que el muflón corso actual desciende de la feralización de las ovejas del Neolítico introducidas en Córcega por el hombre. De nuevo, los taxones endémicos del Pleistoceno se extinguieron durante el Holoceno.[53] Todas las especies corsas modernas se originaron en el continente europeo cercano.[53]
• Chipre

Para Chipre los datos más tempranos provienen del refugio del Holoceno temprano de Akrotiri Aetokremnos, que está considerado por los excavadores como un yacimiento de descuartizamiento de megafauna endémica.[138][153] Akrotiri Aetokremnos proporciona evidencias de la presencia de seres humanos en Chipre hace 9.000 años, y también indicios de la extinción de toda la megafauna endémica y nativa alrededor de ese periodo. Además hay datos del yacimiento pre-cerámico temprano de Shilourokambos descubierto en los 90,[154] fechado entre el final del noveno milenio y la primera mitad del octavo.[53] Para los períodos del Calcolítico y la Edad de Bronce, numerosos análisis fiables han proporcionado datos recientes que atestiguan que la fauna del Neolítico antiguo seguía presente en la isla, junto con algunas nuevas especies (ganado doméstico y équidos, pequeños mamíferos ...).[53] El elefante Palaeoloxodon cypriotes y el hipopótamo Hippopotamus minor, ambas especies endémicas de Chipre,[155] han sido mencionadas en los yacimientos de Akrotiri Aetokremnos, posiblemente asociadas con la ocupación humana más temprana en Chipre, durante el noveno milenio.[53]
• Mallorca
La historia de la relación entre el hombre y los mamíferos en Mallorca muestra el mismo tipo de desarrollo que en las otras grandes islas: las 3 especies endémicas nativas se extinguieron durante el Holoceno tras los primeros asentamientos humanos.[53] Myotragus balearicus es el único megamamífero nativo de una isla mediterránea que sobrevivió a la primera colonización humana. Está bien atestiguado en las capas de Son Matge a lo largo del período de asentamiento temprano, y está claramente ausente de las capas pretalayóticas del mismo yacimiento y de depósitos posteriores.[53]

Entre las especies extintas en las islas mediterráneas están:
- Ciervo enano de Creta (Candiacervus)
- Ciervo gigante de Corcega y Cerdeña Megaloceros cazioti
- Praemegaceros
- Cabra balear de las cavernas (Myotragus balearicus )
- Lirón gigante balear (Hypnomys sp.)
- Lirón gigante de Malta y Sicilia (Leithia sp.)[156]
- Nutria gigante de Cerdeña (Megalenhydris barbaricus)
- Nutria enana de Cerdeña (Sardolutra)
- Perro silvestre de Cerdeña (Cynotherium sardous)
- Mochuelo gigante cretense (Athene cretensis)
- Todos los hipopótamos enanos:
- Hipopótamo enano de Chipre (Hippopotamus minor)
- Hipopótamo enano de Sicilia (Hippopotamus petlandi)
- Hipopótamo enano de Malta (Hippopotamus melitensis)
- Todos los elefantes enanos:
- Palaeoloxodon chaniensis
- Palaeoloxodon cypriotes
- Palaeoloxodon falconeri
- Palaeoloxodon mnaidriensis
Islas de Wrangel y Saint Paul
Wrangel es una isla rusa situada en el océano Ártico que estuvo unida a Beringia hasta hace 12.000 años, y actualmente tiene una superficie de 7680 km².[31] La isla de Saint Paul es la más grande de las islas Pribilof, un grupo de cuatro islas volcánicas de Alaska ubicadas en el Mar de Bering, entre los Estados Unidos y Rusia, tiene unos 100 km². Las islas de Wrangel y Saint Paul son conocidas por ser las últimas masas de tierra que albergaron poblaciones conocidas de mamut lanudo, hace 3.800 y 5000 años respectivamente. Estas fechas contrastan con las dataciones más recientes de mamut lanudo en tierras continentales, fechadas en unos 9.000 años atrás, con lo que hay más de 5 milenios de diferencia entre las fechas de la extinción en el continente y la isla rusa.[31] En el caso de la Isla de Wrangel los primeros yacimientos arqueológicos de Homo sapiens coinciden con los últimos restos de mamut lanudo.[49][79]
Madagascar



La flora y la fauna de Madagascar han seguido una trayectoria evolutiva aislada durante al menos 88 millones de años[157] para producir algunas de las plantas y animales más inusuales en el mundo con niveles extremadamente altos de endemismo.[158] Esta isla continental fue también una de las últimas grandes masas de tierra en ser colonizadas por los seres humanos,hace 2300 años.[159] Al final de un periodo que va de 1400 a 500 años atrás, toda la megafauna endémica de Madagascar había desaparecido. Estos grandes "subfósiles" no eran antepasados de las especies existentes sino que formaban parte de la fauna contemporánea.[160]
Hasta los años 80 del pasado siglo, varios autores defendían que la megafauna de Madagascar había desaparecido por cambios climáticos, alegando que no había evidencias de descuartizamiento en los restos encontrados. A día de hoy ya se han encontrado evidencias confirmadas de que hubo aprovechamiento por parte de los humanos de casi todos los grupos de megafauna —en ocasiones en restos datados inmediatamente después de la llegada del Homo sapiens— como es el caso de los lémures gigantes, los hipopótamos pigmeos o las aves elefante.[63][64] A esta extinción masiva le siguió la conversión del mosaico de bosques y sabanas arboladas en bosques de dosel cerrados y praderas depauperadas de especies.[57] Como consecuencia de la extinción de la megafauna, Madagascar ha perdido comunidades ecológicas enteras,[161] y al menos 34 especies de grandes vertebrados malgaches, entre ellas:
- 3 hipopótamos pigmeos (Hippopotamus)
- Los 17 lémures gigantes conocidos hasta la fecha, pertenecientes a 9 géneros:
- Palaeopropithecus
- Archaeoindris
- Babakotia
- Mesopropithecus
- Megaladapis, el último lémur gigante, Megaladapis edwardsi, fue visto todavía vivo por el gobernador francés, Etienne de Flacourt (Flacourt, 1658)
- Hadropithecus
- Archaeolemur
- Pachylemur
- Daubentonia
- Cerdo hormiguero malgache (Plesiorycteropus)
- Fosa gigante (Cryptoprocta)
- 8 especies de aves elefante pertenecientes a dos géneros:
- Cocodrilo gigante (Voay)
- 2 tortugas gigantes (Aldabrachelys):
Nueva Zelanda

Como en el caso de Madagascar, Nueva Zelanda se separó de Gondwana hace más de 80 millones de años, llevando consigo una fauna y flora únicas y muy arcaicas.[77] La extinción de los moas y resto de megafauna de Nueva Zelanda siguió a la llegada de los seres humanos a finales del siglo XIII y fue una de las últimas extinciones masivas de megafauna del Cuaternario Tardío,[162] siendo el archipiélago de Nueva Zelanda la última gran masa de tierra colonizada por los humanos. Nueva Zelanda albergó un conjunto diverso de megafauna aviar.[77][163][164] La consecuencia de la llegada de los humanos fue la pérdida del 41% de las especies de aves endémicas de las islas.[77] Alrededor de 49 especies de aves (el 51%) de la Isla Norte han desaparecido, junto con 53 especies (47%) de la Isla Sur, y otras 30 están en peligro, de las cuales muchas sobreviven actualmente únicamente porque reciben un manejo activo y costoso.[77] La fauna de Nueva Zelanda que sobrevivió a la extinción masiva lo hizo en buena parte porque las pequeñas islas de la costa proporcionaron el último refugio para muchas especies.[77] Entre las especies extintas se encontraban nueve especies de moa:[165] pájaros gigantes sin alas que variaban en tamaño desde los 12 kg de Euryapteryx curtus a las hembras de unos 250 kg de las dos especies de Dinornis.[77]
Los moas habitaron una variedad de hábitats en todo el archipiélago de Nueva Zelanda hasta su extinción poco después de la llegada de los colonos polinesios, estimada aproximadamente a finales del siglo XIII.[163][77][162] La abundancia de yacimientos arqueológicos bien conservados que contienen pruebas de la explotación a gran escala de moa[166] desecha la controversia del papel de los seres humanos en el evento de extinción, a pesar de lo cual algunos autores aún atribuyen su causa a cambios climáticos, a pesar de que en los últimos 700 años el clima de Nueva Zelanda no ha cambiado.[162] Por otra parte, sí hubo cambios climáticos anteriores sustanciales, que no causaron extinciones.[77] La evidencia de que las aves fueron sacrificadas como alimento es tan abundante que los huesos de algunos yacimientos arqueológicos fueron transportados en tren para ser usados como fertilizantes.[77] La evidencia más sólida de que tras la llegada de los humanos hubo un rápido declive de estas especies es que solo los primeros asentamientos humanos contienen huesos de moa, mientras que ya son raros o inexistentes en los yacimientos de un siglo después al primer asentamiento.[77]
Entre las especies extintas en el evento de extinción del Cuaternario en Nueva Zelanda están las siguientes:


- Águila gigante de Eyles (Circus eylesi)
- Águila gigante de Haast (Harpagornis moorei)
- Todos los moas:
- Moa de los arbustos (Anomalopteryx didiformis)
- Moa de pico ancho de la Isla Norte (Euryapteryx curtus)
- Moa de pico ancho de la Isla Sur (Euryapteryx geranoides)
- Moa crestado (Pachyornis australis)
- Moa elefante (Pachyornis elephantopus)
- Moa de Mappin (Pachyornis mappini)
- Moa oriental (Emeus crassus)
- Moas gigantes del género Dinornis sp.
- Moa gigante de la Isla Sur (Dinornis robustus)
- Moa gigante de la Isla Norte (Dinornis novaezealandiae)
- Moa de pies pesados (Pachyornis elephantopus)
- Moa de las tierras altas (Megalapteryx didinus)
- Ganso gigante de la Isla Norte (Cnemiornis gracilis)
- Ganso gigante de la Isla Sur (Cnemiornis calcitrans)
- Adzebill de la Isla Norte (Aptornis otidiformis)
- Adzebill de la Isla Sur (Aptornis defossor)
- Pato de Finsch (Chenonetta finschi)
- Pato almizclero neozelandés (Biziura delautouri)
- Gallineta de Hodgen (Gallinula hodgenorum)
- Cuervo de Nueva Zelanda (Corvus antipodum)
- Egotelo gigante de Nueva Zelanda (Aegotheles novaezelandiae)
- Takahe de la Isla Norte (Porphyrio mantelli)
- Huia (Heteralocha acutirostris)
- Rascón gigante de Hawkins (Diaphorapteryx hawkinsi)
Otras islas




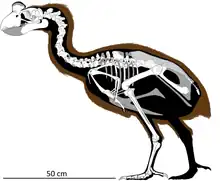
A medida que las técnicas de navegación se hicieron más eficientes y permitieron alcanzar todas las islas del planeta, numerosas extinciones de megafauna tuvieron lugar en los archipiélagos más remotos. Los registros históricos de los siglos XVII al XIX indican que numerosas especies de tortugas gigantes se extinguieron en muchas islas del Océano Índico occidental y el Pacífico oriental durante este período por causa de la caza excesiva.[170] Igualmente, otras especies de reptiles gigantes y muchas aves no voladoras que habían evolucionado sin depredadores terrestres, fueron víctimas de la caza insostenible y los cambios en los ecosistemas causados por la llegada del hombre a otras islas del Pacífico, primero de los colonizadores polinesios y luego de los europeos. Entre las especies extintas están las siguientes:
- Todas las tortugas gigantes cornudas del género Meiolania (extintas en varias islas del Pacífico entre 3000 y 2000 años atrás, tras la llegada del hombre):[167][168]
- Todas las tortugas gigantes del género Cylindraspis:
- Tortuga gigante de Rodrigues con caparazón de silla de montar Cylindraspis vosmaeri (extinta hacia 1795 por caza excesiva en la isla Rodrigues)[57]
- Tortuga gigante de Rodrigues Cylindraspis peltastes (extinta hacia 1795 por caza excesiva en la isla Rodrigues)[57]
- Tortuga gigante de Mauricio con caparazón de silla de montar Cylindraspis triserrata (extinta por caza excesiva en Mauricio hacia 1725 y en islas adyacentes alrededor de 1845)[171]
- Tortuga gigante de Mauricio Cylindraspis inepta (extinta por caza excesiva en Mauricio hacia 1725 y en islas adyacentes alrededor de 1845)[171]
- Tortuga gigante de Reunión Cylindraspis indica (extinta por caza excesiva en Reunión hacia 1840)[172]
- Tres tortugas gigantes de las Galápagos:
- Tortuga gigante de Pinta Chelonoidis abingdonii (extinta por caza excesiva en la isla de Pinta, su último ejemplar conocido fue Solitario George, muerto en 2012, aunque se está estudiando la existencia de individuos híbridos)[173][174]
- Tortuga gigante de Floreana (Chelonoidis elephantopus extinta 15 años después de la vista de Darwin en 1835)[175]
- Tortuga gigante de Fernandina (Chelonoidis phantastica, extinta en 1906 por un ritmo de caza insostenible)[176][177][175]
- Iguana gigante de Fiyi Lapitiguana impensa (extinta hace 3000 años tras la llegada del hombre)[178]
- Iguana gigante de Tonga Brachylophus gibbonsi (extinta un siglo después de la colonización humana del archipiélago hace 2800 años)[179]
- Todos los cocodrilos terrestres del género Mekosuchus:
- Cocodrilo terrestre de Nueva Caledonia Mekosuchus inexpectatus (extinto hace 4000 años tras la llegada del hombre)[180]
- Cocodrilo terrestre de Vanuatu Mekosuchus kalpokasi (extinto hace 3000 años tras la llegada del hombre)[181]
- Galliforme gigante de Nueva Caledonia Sylviornis neocaledoniae, el mayor galliforme conocido (extinto hace unos 1800 años tras la llegada de los Lapita a Nueva Caledonia)[182]
- Azor gigante de Nueva Caledonia Accipiter efficax (extinto hace unos 1800 años tras la llegada del hombre a Nueva Caledonia)[182]
- Lechuza de Nueva Caledonia Tyto letocarti (extinta hace unos 1800 años tras la llegada del hombre a Nueva Caledonia)[182]
- Rascón gigante de Nueva Caledonia Porphyrio kukwiedei (extinto hace unos 1800 años tras la llegada de los Lapita a Nueva Caledonia)[182]
- Falso talégalo gigante de las islas Fiyi Megavitiornis altirostris (extinto después de la colonización de las islas Fiyi por los seres humanos)[183]
- Todos los talégalos gigantes:
- Tálegalo constructor gigante Megapodius molistructor (extinto hace unos 1800 años tras la llegada de los Lapita a Nueva Caledonia)[182]
- Talégalo consumido Megapodius alimentum (extinto después de la colonización de las islas Fiyi y Tonga por los seres humanos)[183]
- Talégalo perdido Megapodius amissus (extinto después de la colonización de las islas Fiyi por los seres humanos)[183]
- Paloma terrestre de Nueva Caledonia Gallicolumba longitarsus (extinta hace unos 1800 años tras la llegada de los Lapita a Nueva Caledonia)[182]
- Cagú de las tierras bajas Rhynochetos orarius (extinta hace unos 1800 años tras la llegada de los Lapita a Nueva Caledonia)[182]
- Agachadiza gigante de Nueva Caledonia Coenocorypha neocaledonica (extinta hace unos 800 años en Nueva Caledonia)[182]
- Agachadiza de Viti Levu Coenocorypha miratropica (extinto después de la colonización de las islas Fiyi por los seres humanos)[183]
- Paloma gigante de Viti Levu Natunaornis gigoura (extinta después de la colonización de las islas Fiyi por los seres humanos)[184]
- Paloma imperial gigante de la isla Henderson Ducula harrisoni (extinta después de la colonización de la isla Henderson por los seres humanos)[185]
- Tórtola-cuco gigante de las islas Marquesas Macropygia heana (extinta después de la colonización de las islas Marquesas por los seres humanos)[186]
- Paloma apuñalada de la isla Henderson Gallicolumba leonpascoi (extinta después de la colonización de la isla Henderson por los seres humanos)[187]
La supervivencia de la megafauna subsahariana: coevolución e ingenuidad insular
Todas las extinciones masivas de megafauna del Cuaternario tardío parecen haber coincidido en el tiempo con la expansión del ser humano moderno a nuevos continentes e islas, excepto en el caso del África subsahariana. Esta región es el lugar de origen de nuestra especie y de los homínidos, y ha sido con diferencia la menos afectada por el proceso de extinción de grandes animales hasta el SXIX, en que comienzan a extinguirse poblaciones enteras y varias especies de megafauna subsahariana con la llegada de las armas de fuego.[188] Hoy se apunta a que el África subsahariana mantuvo su megafauna gracias a que las especies africanas soportaron al menos desde la aparición del género Australopithecus hace unos 4 millones de años, una larga presencia de homínidos cazadores. Así estas especies de megafauna tuvieron que coevolucionar con especies del género Homo y otros géneros anteriores, y desarrollaron una fuerte respuesta defensiva que les lleva a evitar instintivamente el contacto con los homínidos o a iniciar un ataque ante su presencia.[1]
.JPG.webp)
La megafauna del resto del mundo o bien había estado expuesta a una presión cinegética por parte de humanos primitivos menor que la africana aunque importante[189] —es el caso particular de la megafauna eurosiática/paleártica, cuya extinción masiva fue la más lenta de todas tras la llegada del Homo sapiens—[1] o bien no había cohabitado jamás con homínidos de ningún tipo, por lo que no habría desarrollado ningún mecanismo instintivo para identificarnos como depredadores; es el caso de las Américas, Oceanía, la Antártida y las islas oceánicas de todo el planeta.[6] Este ausencia de respuesta defensiva se conoce como ingenuidad insular o ingenuidad ecológica (island tameless o island naivety en inglés) y se ha registrado como factor principal de extinción en numerosas ocasiones.[190] La ingenuidad insular se observa actualmente en aquellas especies endémicas de islas y continentes de reciente descubrimiento como las Galápagos o el continente de la Antártida.[14][6] Este comportamiento está considerado por los ecólogos como un factor clave de vulnerabilidad de las especies autóctonas frente a la aparición de especies alóctonas.[191] Así por ejemplo, los pingüinos del continente antártico muestran un profundo instinto de evasión frente a especies depredadoras con las que han coevolucionado, como la foca leopardo, pero no frente al ser humano, al que no temen.[192]
_females.jpg.webp)
La enorme diferencia entre la escasa cantidad de puntos de descuartizamiento de elefantes africanos y los abundantes puntos para descuartizar proboscídeos cazados encontrados en otros puntos del globo parece apoyar esta hipótesis. Así en Estados Unidos hay más emplazamientos de descuartizamiento de proboscídeos que los que hay de elefantes en toda África, una masa de tierra que es mucho mayor que Estados Unidos. No solo es destacable la diferencia de tamaño entre África y EE. UU., sino que la presencia de homínidos en África se remonta al menos 100 veces más atrás en el tiempo que el corto lapso cronoestratigráfico de presencia humana en América del Norte. Sin embargo, en África hay menos de una docena de yacimientos de descuartizamiento probables, a pesar de que abarcan un rango de tiempo de coexistencia muchísimo más amplio, desde el Plio-Pleistoceno hasta la actualidad. En contraste en EE. UU. hay al menos 14 yacimientos para un periodo al menos 100 veces más corto de coexistencia.[17][20][21] En Eurasia los restos que demuestran un uso intensivo del mamut lanudo como fuente de alimento son aún más abundantes y dan una idea de hasta qué punto este animal fue cazado a gran escala por el ser humano moderno. Hay más de 70 yacimientos a lo largo y ancho de la llanura rusa, donde se han encontrado poblados enteros que se construyeron esencialmente con huesos de mamuts, donde esos huesos fueron también usados como combustible —y para ello tenían que ser relativamente frescos—, y donde los humanos excavaron grandes trampas en el permafrost, que han conservado restos de carne y huesos de mamut. Esta abundancia de yacimientos contrasta significativamente con los doce yacimientos encontrados en África.[193]
Consecuencias de la extinción del Cuaternario tardío
La extinción de la megafauna en buena parte del planeta durante el Cuaternario tardío tuvo efectos devastadores y catastróficos para los ecosistemas terrestres, que llegan hasta nuestros días y cuyas dimensiones en perdidas de biodiversidad y de ecosistemas solo se están empezando a dilucidar.[194][7][195][196][31][54][197]
Efectos sobre la biodiversidad y capacidad de carga de los ecosistemas


.jpg.webp)
La conversión de los bosques abiertos y de las praderas en mosaico, típicos de gran parte del planeta durante el Pleistoceno, a los bosques más uniformes y grandes pastizales que encontramos hoy se considera actualmente una consecuencia de la extinción masiva de la megafauna.[198][31][195] Tales cambios de hábitat habrían sido perjudiciales también para la distribución y abundancia de herbívoros más pequeños, dependientes de la vegetación rica en nutrientes y espacialmente diversa creada por el impacto de los megaherbívoros.[195] Así con la extinción de la megafauna, la relación entre las diferentes especies cambió, hasta el punto de que varias especies de plantas se extinguieron, y los animales supervivientes se vieron afectados de múltiples maneras. Tras la extinción de la megafauna se ha comprobado un aumento en la acumulación de la masa vegetal que eventualmente acaba provocando grandes incendios y cambios severos en la relación entre las especies,[198] su diversidad y la capacidad de carga del ecosistema.[199] Tal vez la consecuencia más visible a día de hoy es la relativa ausencia de animales grandes en relación con ecosistemas que conservan la megafauna original, ausencia que sigue patente incluso en áreas que son abandonadas durante miles de años por el hombre tras la extinción de la megafauna local.[196]
A modo de ejemplo se pueden comparar las densidades de población de la megafauna de parques nacionales de Norteamérica sin presencia del ser humano en la actualidad, con las de los parques nacionales del África subsahariana. En Canadá el parque nacional Jasper contiene aproximadamente 1300 wapitis y 150 alces.[200] El parque tiene aproximadamente 11,000 km cuadrados y está muy bien protegido de la caza furtiva. En contraste, el parque nacional Chobe de tamaño muy similar a Jasper, en Botsuana, sufre furtivismo pero sostiene entre muchas otras especies más de 50.000 elefantes. Por su parte el Serengueti es uno de los espacios naturales menos alterados del mundo a pesar del creciente furtivismo que viene sufriendo en las últimas décadas. Es solo 3 veces mayor que Jasper, pero contiene 1,7 millones de ñus, un cuarto de millón de cebras y medio millón de gacelas, además de elefantes, hipopótamos, jirafas o rinocerontes entre muchas otras especies de megafauna.[201]
En cuanto a los depredadores Jasper alberga aproximadamente 110 osos grizzly, 90 osos negros y solo 45 lobos.[202] El Serengueti tiene más de 3.000 leones, 8.000 hienas y 1.000 leopardos.[201] Estos números son aún más reseñables cuando se considera la creciente presión de la caza furtiva sobre la fauna africana. La población de búfalos cafre en el Serengueti era de 65.000 ejemplares en 1969, pero se ha reducido a 16.000 en la actualidad.[201]
Jasper tiene una temporada de crecimiento más corta, con menos intensa luz solar, mientras que el Serengueti y Chobe sufren caza furtiva desde hace cuatro décadas y una sequía estacional severa. Sin embargo, existe una diferencia abismal en la cantidad de animales mayores de 44 kilos que viven en estos parques.
Es muy difícil estimar la cantidad de animales que estarían presentes si Jasper fuera aún un espacio intacto. Un estudio de restos fósiles en Rusia proporciona algunas cifras para las praderas subárticas del norte de Siberia, entonces parte del mismo bioma que Jasper.[203] Las conclusiones de la investigación sugieren aproximadamente 1 mamut por kilómetro cuadrado en uno de los ambientes más fríos de la tierra. También proporciona cifras para algunas otras especies de megafauna. Si se extrapolan las cifras, un territorio del tamaño de Jasper pudo albergar unos 11.000 mamuts, 66.000 caballos y 165.000 ciervos.[203] En Jasper, el papel de los mamuts europeos se vería probablemente dividido entre los mamuts americanos y el mastodonte. El área también debería contener camellos, perezosos gigantes, castores gigantes, leones americanos, jaguares, tigres de dientes de sable, antílopes saiga, lobos gigantes, osos gigantes, guepardos americanos, bisontes gigantes, varias especies de buey almizclero y berrendos.[15]
Efectos en el paisaje y la distribución de los biomas
_(Kruger_National_Park%252C_2002)_02.jpg.webp)

La extinción de la megafauna en la mayor parte del planeta tuvo consecuencias dramáticas para la extensión y equilibrio de biomas y paisajes, que se vieron profundamente alterados y en algunos casos desaparecieron por completo.[31][54][194][197][196][198] Tras la extinción, los bosques abiertos y las praderas en mosaico de tipo sabana que caracterizaban gran parte del planeta durante el Pleistoceno, retrocedieron y dieron lugar a los extensos bosques cerrados y amplios pastizales que encontramos hoy dominando el paisaje en grandes extensiones fuera del África subsahariana.[195][198]
Los grandes herbívoros tienen un impacto muy importante en la estructura de la vegetación, destacando el equilibrio entre los árboles y las gramíneas. Esto es aún más evidente en el caso de los megaherbívoros debido a su práctica inmunidad a la depredación (no humana) cuando son adultos. Al ser apenas regulados por los depredadores, alcanzan altas densidades en las que pueden transformar radicalmente la estructura de la vegetación.[195][198]
Uno de los estudios más completos para documentar el efecto de la megafauna sobre la vegetación fue realizado mediante aviones que sobrevolaron dos ecosistemas similares de sabana africana durante casi 40 años. La única diferencia entre los dos espacios fue que en uno todos los animales mayores de 5 kg habían sido excluidos, mientras que en el otro seguían presentes. La cubierta vegetal aumentó sobre un 9% a lo largo de 36 años en el sistema sin grandes herbívoros, destacando el papel de los elefantes como especie clave.[206] Este estudio demostró que los elefantes son los principales responsables de la caídas de árboles, y que cada elefante puede desarraigar hasta 1500 árboles por año.[207][205] En su ausencia, la materia vegetal comienza a acumularse y proporciona combustible que acaba provocando un aumento de los incendios forestales.[199]
Los datos paleontológicos también indican un aumento de la biomasa leñosa alrededor de la época de las extinciones de megafauna, pero hasta hace pocos años era difícil determinar si el aumento de la biomasa leñosa era la causa de las extinciones o una consecuencia de las mismas. La dificultad se agravaba porque los restos de megafauna son raros, especialmente en los lagos que generalmente archivan los registros de polen del Cuaternario tardío.[58] Sin embargo, recientemente se ha descubierto que las esporas de Sporormiella, un hongo del estiércol, pueden usarse como un indicador de la abundancia de megafauna, ya que en los registros polínicos de los lagos, el declive de Sporormiella sigue al declive de la megafauna: así en América sus esporas abundan en sedimentos glaciales tardíos, después de la extinción de la megafauna americana se vuelven muy raras a través del Holoceno, y aumentan de nuevo con la introducción histórica del ganado doméstico.[208] Estos datos de Sporormiella en América, y otros muy similares del mismo hongo en Australia[54] indican que las extinciones se vieron seguidas por un aumento de la cubierta arbolada y no al revés.[58]
La reducción de la sabana sudamericana

_teeth_at_the_Smilodon_fatalis_at_the_Gustavo_Orc%C3%A9s_Natural_History_Museum%252C_Quito.JPG.webp)
Las sabanas de América del Sur que han llegado hasta nuestros días se encuentran entre los ecosistemas más amenazados del planeta, con tasas de desforestación a menudo más altas que la propia selva amazónica.[210] América del Sur sufrió una drástica pérdida de animales grandes con 59 especies de megafauna sudamericana extintas entre el Pleistoceno tardío y el Holoceno temprano.[27] Todos los megaherbívoros sudamericanos —herbívoros de 1 tonelada o más de peso corporal— se perdieron, (aproximadamente unas 15 especies).[211] La gran mayoría de los herbívoros de 500 kg o más se extinguieron igualmente, con solo algunas especies de tapires (Tapirus spp.) sobreviviendo hasta nuestros días.[58] Hasta hace poco, ha sido difícil atribuir definitivamente los cambios del ecosistema a la ahora extinta megafauna porque se sabía poco sobre su comportamiento.[58] Debido a la drástica naturaleza de las extinciones en América del Sur, el impacto esperable sobre las comunidades vegetales en esta región es potencialmente más grande que en casi cualquier lugar.[58] Durante el Pleistoceno tardío, Sudamérica tenía mayor abundancia de megahervíboros que África,[58] con un predominio de herbívoros muy grandes. En comparación con la megafauna sudamericana, la mayoría de los mamíferos herbívoros de Australia eran relativamente pequeños y los del África subsahariana son de tamaño intermedio.[212]
Existen razones biogeográficas, paleontológicas y experimentales para pensar que estas extinciones de megafauna pueden haber afectado a la cobertura de la superficie arbolada de la sabana.[58] Por ejemplo, si se comparan la cubierta de árboles de la sabana en África (con megafauna) con la de América del Sur (en gran parte sin ella), para un mismo índice de humedad del suelo la sabana africana tiene menos árboles que la sudamericana,[213] lo que puede deberse a la falta de megaherbivoros en América del Sur.[58] Un estudio estadístico de 2015 realizado por los investigadores Faurby y Svenning intentó resolver esta cuestión. Por una parte se tuvo en cuenta que los animales pasteadores pudieron haber tenido un efecto muy diferente sobre la vegetación leñosa que los ramoneadores. Por lo tanto, era importante determinar el grupo al que pertenecían los animales extintos (pasteadores o ramoneadores).[58]
Un trabajo realizado por investigadores de la Universidad Federal de Sergipe también en 2015 sobre la alimentación de la megafauna sudamericana, puso de manifiesto que las diferencias entre las distintas regiones objeto de estudio eran importantes, incluso dentro de los mismos géneros.[214] Este trabajo separó los resultados en cuatro regiones: En la región intertropical brasileña Equus era un pasteador, Stegomastodon y Toxodon tenían una alimentación mixta, y Eremotherium y Palaeolama eran ramoneadores. En la región amazónica Stegomastodon y Toxodon se comportaban como ramoneadores. En el sur de Brasil, Uruguay y el norte de Argentina Equus y Toxodon eran pastoreadores mientras que los alimentadores mixtos incluían todos los demás taxones como Nothrotherium, Stegomastodon, Hippidion, Scelidodon, Scelidotherium, Neosclerocalyptus, Glyptodon, Macrauchenia y Palaeolama. Ninguno de los taxones analizados en esta región eran ramoneadores estrictos. En Ecuador y Perú Equus era herbívoro y Stegomastodon era un consumidor mixto.[214] Por lo tanto, de acuerdo con este trabajo, el único pastoreador estricto era el género Equus que fue excluido de los análisis estadísticos finales de Faurby y Svenning.[58]
Las simulaciones de modelos estadísticos del estudio de Faurby y Svenning predijeron que en presencia de la megafauna, la superficie terrestre de Sudamérica estaría dominada por sabanas más abiertas con bosques menos extensos. En la práctica, para ecorregiones como el Cerrado de Brasil o el bosque seco chiquitano de Bolivia, se planteaba la hipótesis de que la profusión leñosa actual es mucho mayor de lo que sería con la megafauna aún presente, y los niveles totales de biodiversidad probablemente eran similares o superiores en presencia de la megafauna original.[215] Los bosques muy cerrados pueden suponer un mayor estrés para los animales supervivientes, porque de hecho estas especies coevolucionaron siempre con la megafauna, en unos ecosistemas más abiertos.[195] Los resultados del análisis estadístico predijeron así que las sabanas originales probablemente tenían menos biomasa leñosa y los bosques secos de hoy en día habría sido sabanas con los megaherbívoros.
Cambios en las asociaciones vegetales del Medio Oeste norteamericano

En América del Norte, la deglaciación del Pleistoceno-Holoceno coincidió con la llegada del hombre[21] y con unos cambios bióticos masivos,[56] que incluyeron la extinción de decenas de géneros megafaunísticos,[24] la migración de especies y la reorganización de comunidades terrestres,[217] el crecimiento y declive de comunidades vegetales sin análogos modernos,[218] y el aumento de los incendios.[56] Muchas asociaciones de polen fósiles de América del Norte de entre 17000 y 9000 años atrás carecen de análogos modernos, lo que sugiere que las formaciones vegetales de la época eran, en términos compositivos, diferentes a las de hoy en día.[219] Se ha sugerido que estas comunidades vegetales hoy desaparecidas pueden haber estado vinculadas a la megafauna del Pleistoceno.[218][198]
Usando el hongo de estiércol Sporormiella y otros prototipos paleoecológicos del lago Appleman, Indiana, y de varios lugares del estado de Nueva York, un estudio de 2009 estableció que el declive megafaunístico precedió por un estrecho margen de tiempo a crecientes ciclos de incendios forestales y al desarrollo de comunidades vegetales que no tienen análogos modernos. La pérdida de megaherbívoros pudo así haber alterado la estructura y función del ecosistema al liberar a los plantas leñosas comestibles de la presión herbívora, con la consecuente acumulación de materia vegetal combustible.[56] De acuerdo a lo que indican los restos de Sporormiella, las poblaciones megafaunísticas se derrumbaron a lo largo de casi 1000 años, de 14.800 a 13.700 años atrás, mucho antes de las extinciones finales, y durante el período cálido de BøllingAllerød.[56]


En el lago Appleman, Sporormiella era inicialmente abundante, comenzó a declinar hacia 14.800 años atrás, cayó por debajo del 2% en el estrato de 13.700 años, y se mantuvo por debajo del 2% a partir de entonces. Los huesos más recientes de megafauna de América del Norte se agrupan entre 13.000 y 11.500 años atrás,[120] por lo que el declive de Sporormiella aparentemente indica el colapso de la población local y la extinción de especies funcionales, no la extinción final. La declinación de Sporormiella coincidió aproximadamente con el inicio del período cálido Bølling-Allerød, fechado en Groenlandia en 14.690 años atrás.[220]
Las comunidades no análogas de plantas se desarrollaron a partir de los 13.700 años, después de que Sporormiella declinase por debajo del 2%, y fueron indicadas por aumentos en la abundancia de polen de Fraxinus nigra (fresnos negros) y Ostrya / Carpinus (carpes) mientras los niveles de Picea permanecieron altos. Las comunidades no analógas persistieron hasta 11.900 años ka y fueron seguidas por un período de gran abundancia de Pinus (pino), y finalmente por alta abundancia de Quercus (roble), marcando el inicio de bosques caducifolios del Holoceno temprano en el norte de Indiana. Los regímenes de incendios en Appleman tuvieron tres etapas: Antes de los 14.300 años, el carbón era poco común, indicando pocos o ningún incendio local o una biomasa insuficiente para producir abundante carbón vegetal.[56] Después de 14.300 años , los picos de carbón fueron intermitentes. El carbón vegetal alcanzó por primera vez un pico en el nivel de 14.100 años, durante el declive de Sporormiella y antes del aumento de las abundancias de polen de árboles caducifolios. Esta etapa persistió hasta el estrato de los 10.700 años, cuando las tasas de depósito de carbón y abundancia de polen de Quercus aumentaron rápidamente.[56] El estrecho vínculo entre Sporormiella, los cambios en la vegetación y el fuego en el lago Appleman también aparecen en los resultados de los lugares estudiados en el estado de Nueva York.[56]
De los resultados del estudio se deduce que los impactos humanos fueron una causa plausible: los huesos de mamut con marcas de corte extraídos en el sudeste de Wisconsin datan la presencia humana en la región entre 14.800 y 14.100 años atrás, coetáneos por tanto al declive de Sporormiella en el lago Appleman. Por el contrario, en todos los yacimientos estudiados, el declive de Sporormiella es sustancialmente anterior a la iniciación del Dryas Reciente, descartando el enfriamiento climático abrupto como causa, así como la de un hipotético impacto extraterrestre hace 12.900 años.[56]
El reemplazo de la selva mixta australiana por bosques de esclerófilas

Un reciente estudio de 2012 utilizó un registro ambiental de alta resolución de los últimos 130.000 años para ayudar a resolver la causa de la extinción de la megafauna de Australia, y dilucidar sus consecuencias ecológicas. Los investigadores estudiaron el polen y el carbón para reconstruir la secuencia temporal de cambios en la vegetación, los periodos de incendios y los cambios climáticos, y las esporas del hongo Sporormiella para cuantificar la actividad de los grandes herbívoros.[221] Los resultados del registro de polen fósil, Sporormiella y de restos arqueológicos sugieren que fue la llegada humana en lugar del clima, que permaneció estable, la que causó la extinción de la megafauna.[54] Esta extinción provocó el reemplazo de una selva mixta por vegetación esclerófila a través de una combinación de efectos directos sobre la vegetación tras liberarse de la presión herbívora, y sobre el paisaje tras el aumento de los incendios forestales.[54] Este desplazamiento del ecosistema fue tan grande como cualquier efecto que pudiera tener el cambio climático miles de años después en el planeta, como el del final del último ciclo glacial, e indica la magnitud de los cambios que pueden haber ocurrido después de la extinción de la megafauna en otras partes del mundo durante ese periodo.[54]
Un segundo estudio de 2013[55] llegó a conclusiones similares. La extinción masiva de la megafauna ocurrió en Australia entre 50.000 y 45.000 años atrás,[83][75] un período que coincide con la colonización humana de Australia.[222] También ocurrieron alrededor de este periodo grandes cambios en la vegetación, pero hasta esta década no estaba claro si los cambios de vegetación fueron impulsados por el uso humano del fuego[223] —y por lo tanto contribuyeron al evento de extinción— o si estos cambios en la vegetación fueron una consecuencia de la pérdida de los ramoneadores megafaunísticos.[224][54] Los autores del estudio reconstruyeron los cambios de vegetación ocurridos en el sureste de Australia utilizando la composición isotópica de carbono estable en n-alcanos de la planta Hoya carnosa[225] y los niveles de combustión de biomasa a partir de las tasas de acumulación del biomarcador levoglucosano de un núcleo de sedimento datado de la cuenca Murray-Darling.[55] Los resultados mostraron que de 58.000 a 44.000 años atrás, la abundancia de plantas con la vía de fijación de carbono C4 era generalmente alta, entre 60 y 70%. Hace 43.000 años, la abundancia de plantas C4 cayó al 30% y la quema de biomasa aumentó. Este cambio transitorio comenzó después del período de llegada de los humanos, duró cerca de 3.000 años y siguió directamente a la extinción de la megafauna entre 48.900 y 43.600 años atrás.[83] El estudio indicaba que el desplazamiento de la vegetación no fue la causa de la extinción megafaunística en esta región. En cambio, los datos concordaban con la hipótesis según la cual el cambio de vegetación fue la consecuencia de la extinción de grandes ramoneadores y llevó a la acumulación de vegetación propensa a incendios en el paisaje australiano.[55]
La transformación de la estepa del mamut en la tundra

En 1995 Andrey Zimov y otros cinco investigadores rusos y norteamericanos publicaron un extenso trabajo conjunto sobre las causas de la transformación de la estepa ártica —el mayor bioma terrestre en el Pleistoceno, también conocido como la estepa del mamut por ser este su especie clave—, en la tundra actual.[31] Los modelos climáticos generales muestran que las condiciones ambientales de Beringia durante el Pleistoceno eran más frías que en la actualidad, pero con patrones de viento y precipitaciones en general similares a los actuales.[31] Estas y otras observaciones sugieren que la vegetación esteparia y los suelos secos de Beringia en el Pleistoceno tardío no eran consecuencia directa de un macroclima árido. El pisoteo y el pastoreo de los herbívoros garantizaban la sustitución de musgos y líquenes por gramíneas.[31][195] Las hierbas reducen la humedad del suelo de forma más eficiente que los musgos gracias a unas mayores tasas de evapotranspiración.[31]
Los resultados del modelo de simulación empleado en el estudio, basado en la competencia de las plantas por el agua y la luz, y su sensibilidad a los herbívoros y al suministro de nutrientes, predijeron que cualquiera de los dos tipos de vegetación, una estepa dominada por gramíneas y Artemisia, o una tundra dominada por musgos, pudo haberse desarrollado en Beringia bajo ambos climas, el actual y el del Pleistoceno.[31] El modelo sugiere que la tundra dominada por musgos se ve favorecida cuando el pastoreo se reduce a niveles por debajo del equilibrio entre la actividad de los herbívoros, el clima y la vegetación. Estos resultados en su conjunto apuntan a que la megafauna tenía un efecto lo suficientemente grande sobre la vegetación y la humedad del suelo, como para que su extinción pudiera haber contribuido substancialmente a la transformación de la estepa en tundra durante la transición del Pleistoceno al Holoceno.[31]
Efectos en el transporte de nutrientes
La pérdida de megafauna también tuvo efectos importantes sobre la biogeoquímica de nutrientes.[31][226] La megafauna juega un papel esencial en el transporte lateral de nutrientes minerales en los ecosistemas, tendiendo a traslocarlos de las áreas de gran abundancia a aquellas de menor cantidad. Este movimiento se debe al tiempo entre el consumo de los nutrientes y el tiempo en que los liberan a través de la excreción (o, en menor proporción, a través de la descomposición tras su muerte).[227]
Un reciente estudio de 2013[194] analizó este proceso utilizando un nuevo marco matemático que ponderaba este transporte lateral como un proceso de difusión, y demostró que los animales grandes juegan un papel desproporcionadamente grande en la transferencia horizontal de nutrientes a través de paisajes. Los resultados pusieron de manifiesto que en la Cuenca amazónica de Suramérica, esta difusión lateral se redujo en un 98% tras la extinción de la megafauna.[194]
Dado que se cree que la disponibilidad del fósforo limita la productividad en la mayor parte de la región, la disminución de este transporte desde la parte occidental de la cuenca y desde las planicies aluviales debió de impactar significativamente en la ecología de toda la región, y los efectos pueden no haber terminado de alcanzar su límite.[194][7]
Los autores concluían que la actual limitación de fósforo en la cuenca amazónica puede ser parcialmente una reliquia de un ecosistema sin la conectividad funcional que una vez tuvo. Los resultados apuntaban a que las extinciones de la megafauna del Pleistoceno resultaron en grandes y continuas interrupciones del ciclo biogeoquímico terrestre a escalas continentales, y aumentaron la heterogeneidad en la distribución de los nutrientes a nivel mundial.[194]
Efectos en la dispersión de semillas
La extinción de la megafauna tuvo un profundo impacto en la dispersión de semillas[228][229] Muchos árboles que dependen de los megaherbívoros para la dispersión de sus semillas se han extinguido, otras especies todavía están presentes y sin la megafauna que las dispersaban se están extinguiendo lentamente.

Un estudio de 2011[230] concluyó que los elefantes africanos y asiáticos tienen roles potencialmente únicos en sus comunidades como agentes de dispersión de larga distancia para semillas de todos los tamaños, incluyendo algunas demasiado grandes para ser tragadas por cualquier frugívoro alternativo. Este papel está bien documentado para los elefantes africanos del bosque, las selvas del Congo también proporcionan los mejores ejemplos de especies vegetales aparentemente dispersas solo por elefantes.[231] Los rinocerontes selváticos pueden haber desempeñado un papel como dispersores de semillas superpuesto con el de los elefantes en los bosques tropicales asiáticos, antes de su casi extinción reciente, pero no es probable que el siguiente mamífero no rumiante más grande de Asia, el tapir malayo, disperse semillas grandes de grandes frutos.[232]
Un ejemplo de especies muy conocidas que dependían probablemente de la megafauna son la calabaza y el calabacín, ambos de origen americano. El género Cucurbita (calabazas y calabacines) contiene numerosos linajes domesticados. Estaba ampliamente distribuido en el pasado, pero ha disminuido hasta el punto de que varias de las especies progenitoras de los cultivos, son escasas o desconocidas en el medio silvestre. Un reciente estudio de 2015[233] utilizó la secuenciación de alto rendimiento para analizar plastidios completos de 91 muestras de Cucurbita, que comprendían taxones antiguos, modernos silvestres y modernos domésticos. Este análisis evidenció un camino previamente desconocido para la domesticación en el noreste de México y amplias distribuciones arqueológicas de taxones actualmente desconocidos en el medio silvestre. Además, la similitud de secuencias entre las poblaciones silvestres existentes sugerían una fragmentación reciente. Colectivamente, estos resultados apuntaron a descensos de tipo salvaje que coinciden con la domesticación generalizadapero también a un descenso anterior que coincidiría con la desaparición de los grandes herbívoros, un golpe ecológico crítico para las Cucurbita sp. silvestres.[233]

Cucurbita es solo uno más de muchos taxones anacrónicos del Nuevo Mundo, incluyendo el aguacate (Persea americana), el chocolate (Theobroma cacao), el naranjo de Luisiana (Maclura pomifera), la acacia de tres espinas (Gleditsia triacanthos) y el tecomate (Crescentia cujete).[228]
El naranjo de Luisiana produce una fruta de gran tamaño que ningún animal americano come en la actualidad. Para la planta esto supone una enorme pérdida de energía, ya que el propósito de cualquier fruto es dispersar semillas a través de los animales que lo consumen. Como ningún animal moderno lo come, el fruto cae al suelo y las semillas quedan bajo el árbol. De este modo el rango de distribución de estas plantas se contrae, en vez de extenderse. Cuando los europeos registraron por primera vez el naranjo de Luisiana, su área se limitaba a un pequeño territorio en el centro / norte de Texas. Sin embargo, en el registro polínico fósil aparece extendido por todo el continente. De hecho, hoy sabemos que había 7 especies, los otros 6 disminuyeron sus poblaciones hasta la extinción sin la megafauna. Estos árboles de frutas grandes deberían ser mucho más comunes de lo que son hoy debido a la extinción de la megafauna. Las distribuciones de muestras antiguas muestran que algunas especies de Cucurbita se distribuyeron mucho más ampliamente en el pasado.[228]
También hay evidencia de que el contenido de carbono de los bosques disminuyó a medida que los árboles frutales de frutas grandes disminuyeron en abundancia.[234][228] En Sudamérica un estudio estadístico del 2015 ha puesto de manifiesto que decenas de especies de árboles (más de 60) cuyos frutos se cree que eran dispersados por la megafauna, han visto drásticamente reducida su distribución desde la extinción de estos animales.[234]
Efectos sobre las emisiones de metano y el clima
Los herbívoros producen metano como un subproducto de la fermentación en la digestión. Las grandes poblaciones de megafauna herbívora tienen el potencial de contribuir a la concentración atmosférica de metano, un importante gas de efecto invernadero de acción menos duradera pero más intensa que el CO2.[59] Hoy en día, cerca del 20% de las emisiones anuales de metano provienen del ganado doméstico, y se consideran un factor fundamental del efecto invernadero.[235][236] Estudios recientes han puesto de manifiesto que la extinción de los herbívoros de la megafauna causó una reducción drástica del metano atmosférico.[59][237]

Uno de estos estudios estimó en 2010 las emisiones de metano antes de la llegada de los colonos europeos, del bisonte americano que dominaba las Grandes Planicies de Norteamérica. Según los resultados, el exterminio de 30 millones de bisontes causó un decrecimiento de 2.2 Tg/año.[238] Esto supuso un cambio proporcionalmente muy grande para este lapso de tiempo, en la atmósfera del planeta, y es comparativamente mucho menor al que debió ocurrir tras la extinción de la mayor parte de la megafauna americana tras la llegada del hombre.[239][240] Tales cambios en la atmósfera habrían afectado el clima más allá de su impacto en el carbono, pero también habrían impactado en el albedo y la evapotranspiración.[226][10]
Otro estudio examinó el cambio de la concentración de metano en la atmósfera al final del Pleistoceno, después de la extinción de la megafauna de Eurasia y el continente americano. Después de que los primeros humanos llegaron a las Américas, el 73 % de la megafauna en Norteamérica y el 80% en Sudamérica se extinguieron,[239] al mismo tiempo que las enormes poblaciones de mamut lanudo y otros grandes mamíferos del paleártico desaparecían definitivamente en Eurasia. Los resultados de los modelos matemáticos sugieren que esta extinción produjo una caída en la producción de metano de unos 9.6 teragramos por año. Los registros de núcleos de hielo en los polos muestran un brusco descenso de metano justo en esta época.[10] Esto sugiere que la ausencia de emisiones de metano de la megafauna puede haber provocado el abrupto enfriamiento climático conocido como Dryas Reciente, un repentino cambio climático que los autores que apoyaban la hipótesis del cambio climático consideraban como causa de la extinción masiva de megafauna en las Américas y el paleártico, pero que a la luz de las últimas investigaciones, y de los análisis del registro polínico y de Sporormiella, parece que fue consecuencia de la extinción.[10][54][56][237]
Durante el Último Máximo Glacial, la concentración atmosférica de CO2 era de 80-100 ppmv, menor que en los tiempos preindustriales del Holoceno. En ese momento la estepa del mamut era el bioma más extenso de la Tierra.[59] Para estimar el almacenamiento de carbono en todo el bioma, en 2006 se publicó una investigación en la revista Science que analizaba los datos de almacenamiento en suelos de este bioma que persistieron en el permafrost de Siberia y Alaska hasta la actualidad. Los investigadores desarrollaron un modelo que describe la acumulación de carbonos en suelos y en el permafrost. El modelo muestra un aumento lento pero constante del carbono contenido en el suelo a medida que aparece el permafrost. En la trnasición del Pleistoceno al Holoceno, el loess congelado rico en carbono de Europa y el Sur de Siberia se descongeló y perdió la mayor parte de este carbono. El carbono del suelo disminuye cuando la estepa del mamut se transforma en bosques, estepas y tundra.[59] Como resultado de la extinción de la megafauna que mantenía la estepa, más de 5000 tg de carbono se liberaron a la atmósfera, los océanos y otros ecosistemas terrestres. Los resultados del modelo también mostraron que la restauración del ecosistema de la tundra-estepa mejoraría el almacenamiento de carbono del suelo, mientras que proporcionaría otros servicios ecosistémicos importantes.[59]
Estrategias de recuperación de la megafauna y sus ecosistemas
Las extinciones de megafauna del Cuaternario tardío, al haber transcurrido sin reemplazo ecológico, representan el inicio de una degradación trófica aún en curso,[27][24] que ha dado lugar a la pérdida de gremios funcionales enteros y a una relajación del control de la cadenas tróficas descendentes en los ecosistemas actuales. Una alta proporción de los grandes herbívoros que han sobrevivido al Antropoceno[241] han visto reducidos drásticamente su rango de distribución y abundancia, haciéndolos funcionalmente extintos, o han sido reemplazados por el ganado en gran parte de sus rangos históricos.[242][243][244][245]
Dada la importancia ecológica de los grandes herbívoros modernos, el desplome desde el Pleistoceno tardío hasta la actualidad en la diversidad y biomasa de estos animales, es un evento de máxima importancia en la ecología del planeta. Un creciente número de evidencias apoya la hipótesis de que la pérdida de grandes herbívoros afectó severamente a las plantas leñosas y desató cambios de régimen en los biomas de la Tierra.[198] Muchos de los grandes herbívoros que han sobrevivido a la extinción masiva se encuentran actualmente entre las especies más amenazadas, frente a las amenazas combinadas del uso antropogénico de la tierra y el cambio climático.[245] Las consecuencias ecológicas de las extinciones del Cuaternario tardío son, por lo tanto, fundamentales no solo para comprender los cambios de vegetación a lo largo del Holoceno, sino también para el manejo de los ecosistemas en el Antropoceno actual.[198]
Rewilding y restauración ecológica

El rewilding (que vendría a traducirse como reasilvestramiento en español), se está promoviendo como una alternativa a los planteamientos actuales de conservación de la naturaleza.[246] El rewilding es por tanto un nuevo tipo de gestión de la naturaleza que tiene como objetivo restaurar los procesos naturales, tanto ecológicos como evolutivos, con una mínima intervención humana.[247] El interés por esta práctica de conservación está creciendo en las literaturas popular y científica, y el rewilding es objeto de un creciente debate, tanto entre la comunidad científica y profesionales de la conservación, como fuera de estos colectivos. Su uso se está extendiendo a tierras agrícolas abandonadas de Europa, África, Madagascar, Norteamérica, Rusia, Brasil y varias islas oceánicas.[57][246][247][171][248][249][215] En la práctica sin embargo, el rewilding sigue siendo aún una actividad marginal de conservación.[246] Los cuatro proyectos principales de rewilding en 2017 son los de Oostvaardersplassen (Países Bajos), Yellowstone (EE.UU.), el Parque Pleistoceno (Rusia) y las islas de Mauricio.[246] Un objetivo común de todos los proyectos de rewilding es mantener o aumentar la biodiversidad, al mismo tiempo que se reduce el impacto de las intervenciones humanas presentes y pasadas mediante la restauración de especies y procesos ecológicos.[246]
Rewilding es un término plástico que se ha aplicado a diversos conceptos y prácticas, y sus varios significados, objetivos y enfoques se solapan.[250] El término rewilding surgió por primera vez de una colaboración entre el biólogo de conservación Michael Soulé f|Michael Soulé y el ambientalista David Foreman a finales de los años ochenta, que condujeron a la creación del proyecto Wildlands.[250][251]
El primer proyecto de rewilding: la reintroducción del lobo en Yellowstone

En esta versión norteamericana del término, el rewilding se enfoca en proteger áreas centrales grandes y bien conectadas, y en liberar especies clave, sobre todo lobos.[252][253] Esto se conoció como el enfoque de las 3 Cs (áreas centrales, corredores y carnívoros). La experiencia de la reintroducción de lobos en el parque nacional de Yellowstone se considera comúnmente como el ejemplo práctico más emblemático de este enfoque.[246]
El parque nacional de Yellowstone cubre aproximadamente 898.000 hectáreas de hábitats de montaña, pastizales y bosques. Los seres humanos han vivido en la región por lo menos desde hace 11.000 años, y aunque fue declarado un parque nacional en 1872, diversas formas de intervención continuaron. La mayor parte de la fauna precolombina sobrevivió en el Parque, pero los lobos fueron erradicados a principios del siglo XX. A partir de los años 60 se discutió sobre la reintroducción de la especie, debido a las preocupaciones sobre el impacto de las crecientes poblaciones de wapitis Cervus canadensis y finalmente la reintroducción tuvo lugar entre 1995 y 1996. Las investigaciones subsiguientes sobre el crecimiento del álamo temblón, el sauce y el algodoncillo en los últimos años sugieren que los lobos han iniciado una reestructuración de los ecosistemas del norte de Yellowstone a través de un mejor reclutamiento de especies arbóreas.[254][255] Paralelamente a la disminución de la población de wapitis, la población de bisontes ha aumentado en la zona norte. Los lobos pueden estar permitiendo que la población de bisontes aumente a través de una disminución de la competencia interespecífica con una población actualmente menor de wapitis. También se han observado un aumento de la población de castores, probablemente debido, al menos en parte, al resurgimiento de las comunidades de sauce desde la introducción del lobo.[246]
La extinción del Cuaternario como referencia para restaurar los ecosistemas
La ciencia del rewilding se ha centrado en tres bases históricas diferentes: el Pleistoceno, el Holoceno y los nuevos ecosistemas contemporáneos. Las dos primeras suponen reparar los ecosistemas hasta un estadio anterior o intermedio respecto a las extinciones de megafauna del Cuaternario tardío. Por lo tanto la elección de la línea de base tiene diferentes implicaciones para la conservación.[246]

En 2005 Josh Donlan y 11 investigadores más expandieron la visión inicial en un artículo en la revista científica Nature,[257] que se ha considerado tan influyente como controvertido,[246] pidiendo el reasilvestramiento de partes de Norteamérica a través de la sustitución de la megafauna del pleistoceno perdida en el evento de extinción del Cuaternario.[257] Propusieron que la estructura ecológica de los ecosistemas del Pleistoceno, antes de la extinción de la megafauna, debería ser la línea de base apropiada para la restauración de los ecosistemas del planeta. Para restaurar esta línea de base, los autores sugirieron la introducción de sustitutos de especies cazadas hasta la extinción, como el elefante africano y el león en lugar del mastodonte y el león americano respectivamente.
Un planteamiento similar busca la reconstrucción de ecosistemas insulares mediante el reemplazo de taxones extintos en islas, por ejemplo el uso de tortugas gigantes para la dispersión de semillas en islas oceánicas, a día de hoy en marcha en Madagascar, Galápagos, Rodrigues, Seychelles y Mauricio.[256][57][258][249][171][248][259] El animal introducido actúa como un análogo ecológico, también conocido como proxie, para los taxones extinguidos como resultado de los impactos antropogénicos durante el período colonial en las islas.[246]
Los proyectos de reconstrucción de los ecosistemas previos a las extinciones implican una serie de componentes prácticos, como la gestión pasiva, la reintroducción y la sustitución de taxones, algunos de los cuales han sido objeto de críticas.[260] También plantean una serie de preocupaciones políticas, sociales y éticas en las que entran en conflicto con formas más establecidas de gestión ambiental.[246]
El rewilnding en Europa
_antiquus.jpg.webp)

Una segunda aproximación al rewilding se ha desarrollado en Europa, a partir del interés en las redes ecológicas y el pastoreo natural, junto con una exploración de los desafíos planteados a la conservación a medida que los estudios sobre las causas de la extinción de megafauna del Cuaternario tardío y la Paleoecología avanzan. Un enfoque de red ecológica para la planificación y conservación de los espacios naturales surgió de la constatación de que la protección de espacios concretos por sí sola no aseguraría los objetivos de conservación a largo plazo.[264] El rewilding europeo defiende una configuración espacial ecológica coherente, con espacios naturales centrales, corredores para conectarlos, áreas de restauración y zonas tampón para desarrollar paisajes funcionales interconectados.[264] Esta ordenación geográfica es coherente con el concepto de las 3Cs del rewilding norteamericano.

En el modelo europeo se concede mayor importancia al "pastoreo natural": el pastoreo de animales resistentes fuera de un sistema agrícola. Esto se ha vuelto popular en parte como resultado de los cambios en la agricultura y las preocupaciones sobre el impacto negativo en la biodiversidad del despoblamiento rural y del abandono del campo.[265][266] Un interés por los regímenes de pastoreo también fue desencadenado por la influyente hipótesis de Frans Vera sobre la rotación cíclica de la vegetación.[267] Vera postuló que la vegetación natural de las tierras bajas de Europa no era el bosque cerrado que es fundamental para las hipótesis paleoecológicas predominantes en el Holoceno medio europeo. En su lugar, aboga por un "mosaico cambiante" o un paisaje parecido a un parque, donde los grandes herbívoros desempeñaban un papel ecológico esencial al abrir el dosel forestal.[267][268][269]
La hipótesis de Vera ha sido discutida como modelo para los paisajes del Holoceno medio.[270][271][272] El principal problema tanto de la hipótesis como de los artículos que intentan refutarla es que su línea de base son los ecosistemas del Holoceno, cuando toda la megafauna europea de más de una tonelada, y especialmente sus dos proboscídeos, el elefante europeo y el mamut lanudo —que ejercían probablemente la mayor influencia sobre la vegetación—, ya habían desaparecido del área de estudio en el Pleistoceno tardío, durante el primer pulso de extinción del Cuaternario que afecto a Europa central y meridional entre 40.000 y 20.000 años atrás.[1] Así tanto Vera como los autores que cuestionan su hipótesis se centran en el estudio de los bosques de Quercus petraea y Quercus robur que dominaban la mayor parte de Europa en el Holoceno. Para la extinción norteamericana en las mismas latitudes se ha comprobado recientemente que los bosques de Quercus fueron la etapa final de sustitución de la vegetación original, tras el colapso poblacional de la megafauna.[56] Se hacen así necesarios, por tanto, estudios paleoecológicos regionales que desarrollen análisis comparativos entre el declive de los mayores megaherbívoros del Pleistoceno y el registro de polen fósil en Europa, como los ya realizados en Norteamérica y Australia.[56][54]
En cualquier caso la hipótesis de Vera ha ejercido una fuerte influencia sobre la práctica del rewilding en Europa. El rewilding europeo a través del pastoreo natural generalmente se centra en el restablecimiento de un gremio ecológico de grandes herbívoros - ganado vacuno, caballos, jabalí, castor y bisonte europeo - cuyo pastoreo y ramoneo restaurarían o crearían ecosistemas complejos y ricos en especies, en áreas recuperadas o utilizadas anteriormente para agricultura o silvicultura.[273][274] En este caso, el rewilding puede implicar la creación y liberación en el medio salvaje de animales criados en cautiverio, a veces vinculados a prácticas como la desdomesticación, la reconstrucción y la desextinción.[275] En esta fase, la genética, la anatomía y los comportamientos de animales específicos pueden ser tan importanres como su impacto en el paisaje y los ecosistemas.[276]
Oostvaardersplassen, primer proyecto de rewilding europeo
La reserva de Oostvaardersplassen en los Países Bajos ha surgido como una expresión práctica de este enfoque europeo del rewilding y se presenta como un medio para probar la hipótesis de Vera. El pastoreo natural es también un elemento central de Rewilding Europe: una iniciativa continental destinada a repoblar las tierras altas y las zonas marginales de Europa con megafauna.[277][278][279][280]

Oostvaardersplassen es una reserva de 5.406 ha en la provincia de Flevolanda, en los Países Bajos. Oostvaardersplassen es un pólder en las orillas del Markermeer terminado en 1967. Inspirado por el desarrollo ecológico espontáneo de la zona y la observación de que el pastoreo de los gansos salvajes estaba impulsando la dinámica del hábitat.[246] Frans Vera escribió un artículo sobre el potencial para desarrollar un ecosistema que había desaparecido de los Países Bajos hacía mucho tiempo. Trabajando con sus colegas convencieron a las autoridades y al público en general para crear un nuevo ecosistema.[246]
Los grandes herbívoros se introdujeron en el área en 1983 (uro de Heck), 1984 (caballos konik polacos) y 1992 (ciervos rojos) bajo una política de intervención mínima, de modo que los procesos naturales pudieran ocupar una posición central en el manejo y comunicación del ecosistema. Los herbívoros aumentaron rápidamente. Para el año 2000, sus poblaciones comenzaron a aproximarse a la capacidad de carga, y la mortalidad anual aumentó y se hizo más variable. Las grandes mortandades en los inviernos de 2005 y 2010 causaron un debate público y político importante, en particular de los grupos animalistas y de los agricultores. La disputa interna entre las juntas consultivas gubernamentales sobre conservación de la naturaleza y bienestar de los animales, llevó al ministro a establecer una Comisión Internacional sobre la Gestión de Oostvaardersplassen.[246]
_(prehistoric%253B_Siberia.jpg.webp)

Los informes de esta comisión abordan cuestiones específicas del ministro en materia de ecología, bienestar y ordenación, y juntos establecen principios clave para los proyectos de rewilding.[282][283][246] Oostvaardersplassen es un nuevo ecosistema y la primera reserva en Europa donde la reconstrucción de los niveles tróficos y el proceso natural son fundamentales para la gestión, un experimento de conservación de la naturaleza que ha inspirado y desafiado simultáneamentea todos los agentes implicados en la materia, promoviendo el debate científico, público y político.[246][284]
El parque Pleistoceno en Siberia: reconstrucción de la estepa del mamut
La Estación Científica del Noreste (NESS) y el Parque Pleistoceno son instalaciones científicas ubicadas en el norte de Siberia, a 5 km de la ciudad de Chersky (Yakutia).[285]
La Estación Científica del Noreste se estableció en 1977 y se ha convertido en una de las mayores estaciones de investigación del Ártico del mundo.[246] El Parque Pleistoceno es una importante iniciativa que incluye un intento de restaurar el ecosistema de estepa del mamut, que fue dominante en el Ártico en el Pleistoceno tardío.[285] La iniciativa requiere el reemplazo de los actuales ecosistemas improductivos del norte por pastizales altamente productivos, que tienen una alta densidad animal y una alta tasa de biociclado.[286] La base de partida es que durante el Pleistoceno el comportamiento colectivo de millones de herbívoros competitivos mantuvieron los pastizales impidiendo la aparición de la tundra dominada por los musgos.[31]
Los experimentos con reintroducciones de animales se iniciaron en 1988. Actualmente, el Parque Pleistoceno consiste en un área cerrada de 16 kilómetros cuadrados que alberga varios herbívoros: bisonte europeo, buey almizclero, alce, caballos, wapitís siberianos y renos, aunque los bisontes europeos serán sustituidos por bisontes americanos y yaks.[287] El objetivo es aumentar la densidad herbívora hasta que sea suficiente para influir en la vegetación y el suelo. A medida que aumente la densidad animal, se ampliará el límite del cercado. En última instancia, si los herbívoros llegan a ser suficientemente abundantes, existe el objetivo de aclimatar a los tigres siberianos.[203][285][286][288]
Desextinción
Los avances en clonación, ingeniería genética e investigación con células madre, y la capacidad mejorada de recuperar el ADN antiguo, han abierto la posibilidad de la desextinción, es decir, la recreación de especies extintas.[289]

El potencial de desextinción saltó a la escena pública en marzo de 2013 en la conferencia TEDx De Extinction coorganizada por The Long Now Foundation y National Geographic.[276] En posteriores reuniones se consideraron una serie de especies para su desextinción,[276] y se debatieron los criterios para su selección.[276] También se discutieron los pros y los contras de este nuevo enfoque de conservación.[276]
Si bien se han planteado muchas objeciones serias contra la desextinción,[290][291][292][293] existe la sensación de que se trata de un desarrollo tecnológico inevitable y de que las especies extintas serán resucitadas de uno u otro modo en un futuro cercano.[294] Por lo tanto, puede resultar inútil tratar de prevenir este avance científico, especialmente porque existe un potencial de beneficios significativos para la conservación.[276] Sin embargo, se convierte en una obligación para los biólogos de conservación ayudar a guiar los esfuerzos para decidir qué especies son mejores o peores candidatos para la desextinción,[276] con el objeto de proporcionar una visión general y asegurar que las especies más apropiadas son traídas de vuelta.[276][294]
La especie sobre la que se están concentrando más esfuerzos es el mamut lanudo. Ninguna célula ha dado a día de hoy un genoma completo por sí sola —algo poco probable dada la degradación que sufre el ADN a lo largo del tiempo por la radiación natural— pero al analizar diferentes muestras de varios mamuts, los científicos del Proyecto del Genoma del Mamut de la Universidad Estatal de Pensilvania parecen estar cerca de publicar un genoma de mamut lanudo completo.[295] Liderando otro equipo de investigación, Kevin Campbell de la Universidad de Manitoba ha utilizado un fragmento de ADN para replicar la hemoglobina adaptada al frío extremo que los mamuts sintetizaban.[296] En 2017 al menos cuatro equipos independientes de genetistas trabajan ya en la desextinción del mamut lanudo en Harvard (EE. UU.),[297][298] Corea del Sur,[299] Japón,[300] y Yakutia (Rusia).[301]
Otras especies en cuya desextinción se está trabajando actualmente son el león de las cavernas,[302] el bisonte de estepa,[303] la paloma migratoria,[304] el dodo,[305] y el tigre de Tasmania.[306] En España se logró el nacimiento de un bucardo clonado, que murió al poco de venir al mundo.[307] También se están barajando para su clonación a medio plazo el periquito de Carolina,,[305] los moas,[305] y el rinoceronte lanudo.[308]
Referencias
- «Bigger kill than chill: The uneven roles of humans and climate on late Quaternary megafaunal extinctions (PDF Download Available)». ResearchGate (en inglés). Consultado el 28 de febrero de 2017.
- «Megafaunal extinctions and their consequences in the tropical Indo-Pacific (PDF Download Available)». ResearchGate (en inglés). Consultado el 25 de febrero de 2017.
- Milsom, Clare; Rigby, Sue (2009). Fossils at a Glance (2nd edición). John Wiley & Sons. p. 134. ISBN 1405193360.
- History of Life. Universidad de Berkeley.
- Zimmer, Carl (1 de marzo de 2017). «Scientists Say Canadian Bacteria Fossils May Be Earth’s Oldest». The New York Times. ISSN 0362-4331. Consultado el 2 de marzo de 2017.
- «Human Dispersal and Late Quaternary Megafaunal Extinctions: the Role of the Americas in the Global Puzzle (PDF Download Available)». ResearchGate (en inglés). Consultado el 27 de febrero de 2017.
- «The shifted baseline: Prehistoric defaunation in the tropics and its consequences for biodiversity conservation (PDF Download Available)». ResearchGate (en inglés). Consultado el 27 de febrero de 2017.
- «Prehistoric Overkill: The Global Model». ResearchGate (en inglés). Consultado el 28 de febrero de 2017.
- Kolbert, Elizabeth (2014). The Sixth Extinction: An Unnatural History. Bloomsbury Publishing. ISBN 9780805092998.
- «Geophysical Research Letters, Vol. 37, L15703, Biophysical feedbacks between the Pleistocene megafauna extinction and climate: The first human-induced global warming?». Archivado desde el original el 29 de marzo de 2017. Consultado el 3 de abril de 2017.
- «Megafauna Died From Big Kill, Not Big Chill». ResearchGate (en inglés). Consultado el 27 de febrero de 2017.
- «The Impact of Hunting on the Mammalian Fauna of Tropical Asian Forests (PDF Download Available)». ResearchGate (en inglés). Consultado el 27 de febrero de 2017.
- «Exceptional record of mid-Pleistocene vertebrates helps differentiate climatic from anthropogenic ecosystem perturbations (PDF Download Available)». ResearchGate (en inglés). Consultado el 25 de febrero de 2017.
- «Human arrival scenarios have a strong influence on interpretations of the late Quaternary extinctions (PDF Download Available)». ResearchGate (en inglés). Consultado el 21 de febrero de 2017.
- Fiedel, S. (2009). «Sudden Deaths: The Chronology of Terminal Pleistocene Megafaunal Extinction». American Megafaunal Extinctions at the End of the Pleistocene. Vertebrate Paleobiology and Paleoanthropology. pp. 21-37. ISBN 978-1-4020-8792-9. doi:10.1007/978-1-4020-8793-6_2.
- Waters, M. R.; Stafford, T. W. (2007). «Redefining the Age of Clovis: Implications for the Peopling of the Americas». Science 315 (5815): 1122-1126. Bibcode:2007Sci...315.1122W. doi:10.1126/science.1137166.
- Surovell, T. A.; Waguespack, N. M. (15 de noviembre de 2008). «How many elephant kills are 14?: Clovis mammoth and mastodon kills in context». Quaternary International 191 (1): 82-97. Bibcode:2008QuInt.191...82S. doi:10.1016/j.quaint.2007.12.001. Archivado desde el original el 11 de agosto de 2017.
- «Paleoindians, Proboscideans, and Phytotoxins: Exploring the Feasibility of Poison Hunting During the Last Glacial-Interglacial Transition». ResearchGate (en inglés). Consultado el 23 de febrero de 2017.
- «Saving the World's Terrestrial Megafauna (PDF Download Available)». ResearchGate (en inglés). Consultado el 24 de febrero de 2017.
- «The role of mammoths in rapid Clovis dispersal (PDF Download Available)». ResearchGate (en inglés). Consultado el 25 de febrero de 2017.
- «The Early Settlement of North America». Cambridge University Press. Consultado el 25 de febrero de 2017.
- «Environmental Values in a globalising World (PDF Download Available)». ResearchGate (en inglés). Consultado el 25 de febrero de 2017.
- Prescott, Graham W.; Williams, David R.; Balmford, Andrew; Green, Rhys E.; Manica, Andrea (20 de marzo de 2012). «Quantitative global analysis of the role of climate and people in explaining late Quaternary megafaunal extinctions». Proceedings of the National Academy of Sciences (en inglés) 109 (12): 4527-4531. ISSN 0027-8424. PMC 3311372. PMID 22393004. doi:10.1073/pnas.1113875109. Consultado el 2 de marzo de 2017.
- «Late Quaternary Extinctions: State of the Debate (PDF Download Available)». ResearchGate (en inglés). Consultado el 26 de marzo de 2017.
- Saltré, Frédérik; Rodríguez-Rey, Marta; Brook, Barry W.; Johnson, Christopher N; Turney, Chris S. M.; Alroy, John; Cooper, Alan; Beeton, Nicholas et al. (29 de enero de 2016). «Climate change not to blame for late Quaternary megafauna extinctions in Australia». Nature Communications (en inglés) 7. ISSN 2041-1723. PMC 4740174. PMID 26821754. doi:10.1038/ncomms10511. Consultado el 28 de febrero de 2017.
- Surovell, Todd A.; Byrd Finley, Judson; Smith, Geoffrey M.; Brantingham, P. Jeffrey; Kelly, Robert (1 de agosto de 2009). «Correcting temporal frequency distributions for taphonomic bias». Journal of Archaeological Science 36 (8): 1715-1724. doi:10.1016/j.jas.2009.03.029. Consultado el 4 de marzo de 2017.
- Sandom, Christopher; Faurby, Søren; Sandel, Brody; Svenning, Jens-Christian (22 de julio de 2014). «Global late Quaternary megafauna extinctions linked to humans, not climate change». Proc. R. Soc. B (en inglés) 281 (1787): 20133254. ISSN 0962-8452. PMC 4071532. PMID 24898370. doi:10.1098/rspb.2013.3254. Consultado el 2 de marzo de 2017.
- Steadman, David W.; Martin, Paul S.; MacPhee, Ross D. E.; Jull, A. J. T.; McDonald, H. Gregory; Woods, Charles A.; Iturralde-Vinent, Manuel; Hodgins, Gregory W. L. (16 de agosto de 2005). «Asynchronous extinction of late Quaternary sloths on continents and islands». Proceedings of the National Academy of Sciences of the United States of America 102 (33): 11763-11768. ISSN 0027-8424. PMC 1187974. PMID 16085711. doi:10.1073/pnas.0502777102. Consultado el 10 de marzo de 2017.
- «Quaternary Extinctions: A Prehistoric Revolution (PDF Download Available)». ResearchGate (en inglés). Consultado el 28 de febrero de 2017.
- McGlone, M. (23 de marzo de 2012). «The Hunters Did It». Science 335 (6075): 1452-1453. doi:10.1126/science.1220176. Consultado el 25 de marzo de 2012.
- Zimov, S.A. et al. (1994). «Steppe-tundra transition: a herbivore-driven biome shift at the end of the pleistocene.». American Naturalist. 146:765-794.
- Edmeades, Baz. «Megafauna — First Victims of the Human-Caused Extinction». (internet-published book with Foreword by Paul S. Martin). Archivado desde el original el 2 de abril de 2001. Consultado el 4 de octubre de 2010.
- Araujo, B. (2013). «Pleistocene-Holocene extinctions: distinguishing between climatic and anthropic causes.». Universidade federal do Río de Janeiro.
- «Selective hunting of juveniles as a cause of the imperceptible overkill of the Australian Pleistocene 'megafauna' (PDF Download Available)». ResearchGate (en inglés). Consultado el 8 de marzo de 2017.
- Martin, P.S. (1967). «Pleistocene overkill». Natural History 76: 32-38.
- «Updating Martin's global extinction model (PDF Download Available)». ResearchGate (en inglés). Consultado el 28 de febrero de 2017.
- «Megafaunal extinctions and their consequences in the tropical Indo-Pacific (disponible descarga en PDF)». ResearchGate (en inglés). Consultado el 25 de febrero de 2017.
- «Turtles and Tortoises of the World During the Rise and Global Spread of Humanity: First Checklist and Review of Extinct Pleistocene and Holocene Chelonians. (PDF Download Available)». ResearchGate (en inglés). Consultado el 7 de marzo de 2017.
- «The nature of megafaunal extinctions during the MIS 3–2 transition in Japan (PDF Download Available)». ResearchGate (en inglés). Consultado el 12 de marzo de 2017.
- «Island-wide aridity did not trigger recent megafaunal extinctions in Madagascar (PDF Download Available)». ResearchGate (en inglés). Consultado el 17 de marzo de 2017.
- «A premature burial: Comments on Grayson and Meltzer's "Requiem for overkill" (PDF Download Available)». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- «Competition, Predation, and the Evolution and Extinction of Steller’s Sea Cow, Hydrodamalis Gigas». Marine Mammal Science (Society for Marine Mammalogy) 11 (3): 391-4. julio de 1995. doi:10.1111/j.1748-7692.1995.tb00294.x. Archivado desde el original el 11 de mayo de 2011. Consultado el 21 de febrero de 2017.
- «Nineteenth-century explanations of Pleistocene extinctions: A review and analysis». ResearchGate (en inglés). Consultado el 28 de febrero de 2017.
- «The ghosts of mammals past: Biological and geographical patterns of global mammalian extinction across the holocene». ResearchGate (en inglés). Consultado el 28 de febrero de 2017.
- «Assessing the Causes of Late Pleistocene Extinctions on the Continents (PDF Download Available)». ResearchGate (en inglés). Consultado el 28 de febrero de 2017.
- «The Deglaciation of the Northern Hemisphere: A Global Perspective». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- «Late Pleistocene Megafaunal Extinctions». ResearchGate (en inglés). Consultado el 21 de febrero de 2017.
- «Human Dispersal and Late Quaternary Megafaunal Extinctions: the Role of the Americas in the Global Puzzle (PDF Download Available)». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- «Holocene dwarf mammoths from Wrangel Island in the Siberian Arctic». ResearchGate (en inglés). Consultado el 26 de marzo de 2017.
- T.W. Stafford Jr, V.S. Sellars, J.R. Johnson (March 20–24, 2002.). «Chronostratigraphy at Arlington Springs, a Paleoindian site in insular California». Paper presented at 67th annual meeting of Society for American Archaeology, Denver.
- L.D. Agenbroad (March 20–24, 2002.). «Pygmy mammoths (Mammuthus exilis) and a human presence on Santa Rosa Island: coincidence or extinction?». Paper presented at 67th Annual Meeting of Society for American Archaeology, Denver.
- M. Masseti (2001). «Did endemic dwarf elephants survive on Mediterranean islands up to protohistorical times?». The World of Elephants - International Congress, Rome.
- «The Large "True" Mediterranean Islands as a Model for The Holocene Human Impact on the European Vertebrate Fauna? Recent Data and New Reflections (PDF Download Available)». ResearchGate (en inglés). Consultado el 12 de marzo de 2017.
- Rule, S.; Brook, B. W.; Haberle, S. G.; Turney, C. S. M.; Kershaw, A. P. (23 de marzo de 2012). «The Aftermath of Megafaunal Extinction: Ecosystem Transformation in Pleistocene Australia». Science 335 (6075): 1483-1486. doi:10.1126/science.1214261. Consultado el 25 de marzo de 2012.
- Lopes dos Santos, Raquel A.; De Deckker, Patrick; Hopmans, Ellen C.; Magee, John W.; Mets, Anchelique; Sinninghe Damsté, Jaap S.; Schouten, Stefan (1 de agosto de 2013). «Abrupt vegetation change after the Late Quaternary megafaunal extinction in southeastern Australia». Nature Geoscience (en inglés) 6 (8): 627-631. ISSN 1752-0894. doi:10.1038/ngeo1856. Consultado el 24 de marzo de 2017.
- «Pleistocene Megafaunal Collapse, Novel Plant Communities, and Enhanced Fire Regimes in North America (PDF Download Available)». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- «Using a surviving lineage of Madagascar's vanished megafauna for ecological restoration (PDF Download Available)». ResearchGate (en inglés). Consultado el 17 de marzo de 2017.
- «The impact of the megafauna extinctions on savanna woody cover in South America (PDF Download Available)». ResearchGate (en inglés). Consultado el 18 de marzo de 2017.
- Walter, K. M.; Zimov, S. A.; Chanton, J. P.; Verbyla, D.; Chapin, F. S. (7 de septiembre de 2006). «Methane bubbling from Siberian thaw lakes as a positive feedback to climate warming». Nature (en inglés) 443 (7107): 71-75. ISSN 0028-0836. doi:10.1038/nature05040. Consultado el 28 de marzo de 2017.
- «Timing of Quaternary megafaunal extinction in South America in relation to human arrival and climate change (PDF Download Available)». ResearchGate (en inglés). Consultado el 28 de febrero de 2017.
- «Combination of humans, climate, and vegetation change triggered Late Quaternary megafauna extinction in the Última Esperanza region, southern Patagonia, Chile (PDF Download Available)». ResearchGate (en inglés). Consultado el 28 de febrero de 2017.
- «The Associational Critique of Quaternary Overkill and Why It is Largely Irrelevant to the Extinction Debate (disponible descarga en PDF)». ResearchGate (en inglés). Consultado el 4 de marzo de 2017.
- «Evidence of butchery of giant lemurs in Madagascar (PDF Download Available)». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- «Box 4. Butchered sloth lemurs». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- «Cultural innovation and megafauna interaction in the early settlement of arid Australia (PDF Download Available)». ResearchGate (en inglés). Consultado el 28 de marzo de 2017.
- «The exploitation of megafauna during the earliest peopling of the Americas: An examination of nineteenth-century fossil collections». ResearchGate (en inglés). Consultado el 28 de marzo de 2017.
- Grayson, D. K.; Meltzer, D. J. (2002). «Clovis Hunting and Large Mammal Extinction: A Critical Review of the Evidence». Journal of World Prehistory 16 (4): 313. doi:10.1023/A:1022912030020.
- Surovell, T. A.; Waguespack, N. M. (15 de noviembre de 2008). «How many elephant kills are 14?: Clovis mammoth and mastodon kills in context». Quaternary International 191 (1): 82-97. Bibcode:2008QuInt.191...82S. doi:10.1016/j.quaint.2007.12.001. Archivado desde el original el 11 de agosto de 2017.
- Andersen, K. K.; Azuma, N.; Barnola, J.-M.; Bigler, M.; Biscaye, P.; Caillon, N.; Chappellaz, J.; Clausen, H. B. et al. (9 de septiembre de 2004). «High-resolution record of Northern Hemisphere climate extending into the last interglacial period». Nature (en inglés) 431 (7005): 147-151. ISSN 0028-0836. doi:10.1038/nature02805. Consultado el 28 de marzo de 2017.
- Walker, Mike (2005). Department of Archaeology and Anthropology University of Wales, Lampeter, UK, ed. Quaternary Dating Methods. John Wiley and Sons Lld. Archivado desde el original el 14 de julio de 2014.
- Mead, J.I., Meltzer, D.J. (1984). «North American late Quaternary extinctions and the radiocarbon record.». Martin, P.S., Klein, R.G. (Eds.) Quaternary Extinctions: a Prehistoric Revolution. Univ. Arizona Press, Tucson, pp. 441e450.
- «A chronology for late prehistoric Madagascar (PDF Download Available)». ResearchGate (en inglés). Consultado el 16 de marzo de 2017.
- Iwase, Akira; Hashizume, Jun; Izuho, Masami; Takahashi, Keiichi; Sato, Hiroyuki (26 de marzo de 2012). «Timing of megafaunal extinction in the late Late Pleistocene on the Japanese Archipelago». Quaternary International. Mammoths and Their Relatives 1: Biotopes, Evolution and Human Impact V International Conference, Le Puy-en-Velay, 2010 255: 114-124. doi:10.1016/j.quaint.2011.03.029. Consultado el 16 de marzo de 2017.
- Grün, R.; Wells, R.; Eggins, S.; Spooner, N.; Aubert, M.; Brown, L.; Rhodes, E. (1 de agosto de 2008). «Electron spin resonance dating of South Australian megafauna sites». Australian Journal of Earth Sciences 55 (6-7): 917-935. ISSN 0812-0099. doi:10.1080/08120090802097419. Consultado el 16 de marzo de 2017.
- «ESR and U-Series analyses of faunal material from Cuddie Springs, NSW, Australia: implications for the timing of the extinction of the Australian megafauna». ResearchGate (en inglés). Consultado el 16 de marzo de 2017.
- Stringer, C. and Andrew, P. (2005). The Complete World of Human Evolution. Londres: Thames & Hudson.
- «The Lost World of the Moa: Prehistoric Life of New Zealand, Trevor H Worthy & Richard N Holdaway (PDF Download Available)». ResearchGate (en inglés). Consultado el 18 de marzo de 2017.
- Todd A. Surovell, Nicole M. Waguespack (2008). «How many elephant kills are 14? Clovis mammoth and mastodon kills in context». Quaternary International 191 (2008) 82–97. Archivado desde el original el 11 de agosto de 2017.
- Lister, A. M. (25 de marzo de 1993). «Mammoths in miniature». Nature (en inglés) 362 (6418): 288-289. doi:10.1038/362288a0. Consultado el 26 de marzo de 2017.
- «Australia's Mammal Extinctions: A 50,000 Year History». ResearchGate (en inglés). Consultado el 28 de febrero de 2017.
- «A quantitative model for distinguishing between climate change, human impact, and their synergistic interactions as drivers of the late Quaternary megafaunal extinctions (PDF Download Available)». ResearchGate (en inglés). Consultado el 28 de febrero de 2017.
- «Colonization of the Americas, ‘Little Ice Age’ climate, and bomb-produced carbon: Their role in defining the Anthropocene (PDF Download Available)». ResearchGate (en inglés). Consultado el 28 de febrero de 2017.
- Roberts, Richard G.; Flannery, Timothy F.; Ayliffe, Linda K.; Yoshida, Hiroyuki; Olley, Jon M.; Prideaux, Gavin J.; Laslett, Geoff M.; Baynes, Alexander et al. (8 de junio de 2001). «New Ages for the Last Australian Megafauna: Continent-Wide Extinction About 46 000 Years Ago». Science (en inglés) 292 (5523): 1888-1892. ISSN 0036-8075. PMID 11397939. doi:10.1126/science.1060264. Consultado el 4 de marzo de 2017.
- «Megafaunal extinctions and their consequences in the tropical Indo-Pacific (disponible descarga en PDF)». ResearchGate (en inglés). Consultado el 25 de febrero de 2017.
- LLC, Revolvy,. «"Megalochelys atlas" on Revolvy.com». www.revolvy.com (en inglés). Consultado el 29 de marzo de 2017.
- «Prehistoric Mammals of Australian and New Guinea - 100 Million Years of Evolution». ResearchGate (en inglés). Consultado el 25 de febrero de 2017.
- «Characteristics of Pleistocene megafauna extinctions in Southeast Asia (disponible descarga en PDF)». ResearchGate (en inglés). Consultado el 25 de febrero de 2017.
- «Mammoth and other proboscideans in China during the Late Pleistocene». ResearchGate (en inglés). Consultado el 29 de marzo de 2017.
- «Discovery of Elephas cf. namadicus from the Late Pleistocene Strata of Marginal Ganga Plain (PDF Download Available)». ResearchGate (en inglés). Consultado el 29 de marzo de 2017.
- «Characteristics of Pleistocene megafauna extinctions in Southeast Asia (PDF Download Available)». ResearchGate (en inglés). Consultado el 29 de marzo de 2017.
- «A review of evidence for postulated Middle Pleistocene occupations in Viet Nam». ResearchGate (en inglés). Consultado el 29 de marzo de 2017.
- «On the complex of the Stegodon-Ailuropoda fauna of southern China and the chronological position of Gigantopithecus blacki V. Koenigswald». ResearchGate (en inglés). Consultado el 29 de marzo de 2017.
- «Pleistocene mammalian faunas of China.». Pleistocene mammalian faunas of China. In: Wu, R., Olsen, J. (Eds.), Palaeoanthropology and Palaeolithic Archaeology in the People's Republic of China. Academic Press, Orlando, pp. 267–289. 1985.
- «The Impact of Hunting on the Mammalian Fauna of Tropical Asian Forests (disponible descarga en PDF)». ResearchGate (en inglés). Consultado el 25 de febrero de 2017.
- «Prehistoric Periods in Japan». www.t-net.ne.jp. Consultado el 12 de marzo de 2017.
- Clarkson, Chris; Smith, Mike; Marwick, Ben; Fullagar, Richard; Wallis, Lynley A.; Faulkner, Patrick; Manne, Tiina; Hayes, Elspeth et al. (1 de junio de 2015). «The archaeology, chronology and stratigraphy of Madjedbebe (Malakunanja II): A site in northern Australia with early occupation». Journal of Human Evolution 83: 46-64. doi:10.1016/j.jhevol.2015.03.014. Consultado el 4 de marzo de 2017.
- «Both half right: Updating the evidence for dating first human arrivals in Sahul (disponible descarga en PDF)». ResearchGate (en inglés). Consultado el 4 de marzo de 2017.
- Bird, Michael I.; Hutley, Lindsay B.; Lawes, Michael J.; Lloyd, Jon; Luly, Jon G.; Ridd, Peter V.; Roberts, Richard G.; Ulm, Sean et al. (1 de julio de 2013). «Humans, megafauna and environmental change in tropical Australia». Journal of Quaternary Science (en inglés) 28 (5): 439-452. ISSN 1099-1417. doi:10.1002/jqs.2639. Consultado el 4 de marzo de 2017.
- «The human colonisation of Australia: optical dates of 53,000 and 60,000 years bracket human arrival at Deaf Adder Gorge, Northern Territory». ResearchGate (en inglés). Consultado el 4 de marzo de 2017.
- «The Archaeology of Australia's Deserts». Cambridge University Press (en inglés). Consultado el 4 de marzo de 2017.
- Johnson, Christopher N.; Brook, Barry W. (22 de diciembre de 2011). «Reconstructing the dynamics of ancient human populations from radiocarbon dates: 10 000 years of population growth in Australia». Proc. R. Soc. B (en inglés) 278 (1725): 3.748-3.754. ISSN 0962-8452. PMC 3203495. PMID 21561972. doi:10.1098/rspb.2011.0343. Consultado el 4 de marzo de 2017.
- van der Kaars, Sander; Miller, Gifford H.; Turney, Chris S. M.; Cook, Ellyn J.; Nürnberg, Dirk; Schönfeld, Joachim; Kershaw, A. Peter; Lehman, Scott J. (20 de enero de 2017). «Humans rather than climate the primary cause of Pleistocene megafaunal extinction in Australia». Nature Communications (en inglés) 8. ISSN 2041-1723. PMC 5263868. PMID 28106043. doi:10.1038/ncomms14142. Consultado el 8 de marzo de 2017.
- «Geological Implications of Impacts of Large Asteroids and Comets on the Earth». ResearchGate (en inglés). Consultado el 5 de marzo de 2017.
- «Late Pleistocene Australian Marsupial DNA Clarifies the Affinities of Extinct Megafaunal Kangaroos and Wallabies (PDF Download Available)». ResearchGate (en inglés). Consultado el 29 de marzo de 2017.
- «Modern Examples of Extinctions (PDF Download Available)». ResearchGate (en inglés). Consultado el 29 de marzo de 2017.
- Extinctions in Near Time: Causes, Contexts, and Consequences. Springer Science+Business Media, LLC. 1999. ISBN 978-1-4757-5202-1.
- «The ghosts of mammals past: Biological and geographical patterns of global mammalian extinction across the holocene». ResearchGate (en inglés). Consultado el 27 de febrero de 2017.
- Pitulko, V. V.; Nikolsky, P. A.; Girya, E. Yu; Basilyan, A. E.; Tumskoy, V. E.; Koulakov, S. A.; Astakhov, S. N.; Pavlova, E. Yu et al. (2 de enero de 2004). «The Yana RHS Site: Humans in the Arctic Before the Last Glacial Maximum». Science (en inglés) 303 (5654): 52-56. ISSN 0036-8075. PMID 14704419. doi:10.1126/science.1085219. Consultado el 4 de marzo de 2017.
- «Global late Quaternary megafauna extinctions linked to humans, not climate change (disponible descarga en PDF)». ResearchGate (en inglés). Consultado el 4 de marzo de 2017.
- «The Complete World of Human Evolution by Chris Stringer and Peter Andrews». New Scientist (en inglés estadounidense). Consultado el 4 de marzo de 2017.
- Elias, Scott; Mock, Cary (25 de marzo de 2013). Encyclopedia of Quaternary Science (en inglés). Newnes. ISBN 9780444536426.
- «Hunting the Extinct Steppe Bison (Bison priscus) Mitochondrial Genome in the Trois-Frères Paleolithic Painted Cave (PDF Download Available)». ResearchGate (en inglés). Consultado el 29 de marzo de 2017.
- «Early cave art and ancient DNA record the origin of European bison (PDF Download Available)». ResearchGate (en inglés). Consultado el 29 de marzo de 2017.
- «Tracking of wisent–bison–yak mitochondrial evolution (PDF Download Available)». ResearchGate (en inglés). Consultado el 29 de marzo de 2017.
- «The Recently Extinct Plants and Animals Database cubit: The Recently Extinct Plants and Animals Database Extinct Mammals: Placental Mammals: Spirocerus kiakhtensis». cubits.org. Archivado desde el original el 9 de septiembre de 2017. Consultado el 6 de marzo de 2017.
- «Habitat conditions for Camelus knoblochi and factors in its extinction by Vadim V. Titov».
- Ghezzo, Elena; Boscaini, Alberto; Madurell-Malapeira, Joan; Rook, Lorenzo (16 de diciembre de 2014). «Lynx remains from the Pleistocene of Valdemino cave (Savona, Northwestern Italy), and the oldest occurrence of Lynx spelaeus (Carnivora, Felidae)». Rendiconti Lincei (en inglés) 26 (2): 87-95. ISSN 2037-4631. doi:10.1007/s12210-014-0363-4.
- Münzel, Susanne C.; Rivals, Florent; Pacher, Martina; Döppes, Doris; Rabeder, Gernot; Conard, Nicholas J.; Bocherens, Hervé (7 de agosto de 2014). «Behavioural ecology of Late Pleistocene bears (Ursus spelaeus, Ursus ingressus): Insight from stable isotopes (C, N, O) and tooth microwear». Quaternary International. Fossil remains in karst and their role in reconstructing Quaternary paleoclimate and paleoenvironments. 339–340: 148-163. doi:10.1016/j.quaint.2013.10.020.
- «The timing of ostrich existence in Central Asia: AMS 14C age of eggshells from Mongolia and southern Siberia (a pilot study) (PDF Download Available)». ResearchGate (en inglés). Consultado el 18 de marzo de 2017.
- Fiedel, Stuart; Haynes, Gary (1 de enero de 2004). «A premature burial: comments on Grayson and Meltzer's “Requiem for overkill”». Journal of Archaeological Science 31 (1): 121-131. doi:10.1016/j.jas.2003.06.004. Consultado el 27 de marzo de 2017.
- «Post-Clovis survival of American Mastodon in the southern Great Lakes Region of North America (PDF Download Available)». ResearchGate. Consultado el 11 de marzo de 2017.
- «Whos who in the Pleistocene: A mammalian bestiary». ResearchGate. Consultado el 11 de marzo de 2017.
- Todd A. Surovell, Nicole M. Waguespack (2008). «How many elephant kills are 14? Clovis mammoth and mastodon kills in context». Quaternary International 191 (2008) 82–97.
- Todd A. Surovell, Nicole M. Waguespack (2008). «How many elephant kills are 14? Clovis mammoth and mastodon kills in context». / Quaternary International 191 (2008) 82–97. Archivado desde el original el 11 de agosto de 2017.
- «A Cladistic Analysis among Trilophodont gomphotheres (Mammalia, Proboscidea) with special attention to the South American genera (PDF Download Available)». ResearchGate (en inglés). Consultado el 26 de marzo de 2017.
- «Fossilworks: Agalmaceros». fossilworks.org. Consultado el 24 de enero de 2016.
- «Fossilworks: Agalmaceros blicki». fossilworks.org. Consultado el 24 de enero de 2016.
- «Fossilworks: Valgipes». fossilworks.org. Consultado el 24 de enero de 2016.
- Martin, Paul Schultz (1 de enero de 2005). Twilight of the Mammoths: Ice Age Extinctions and the Rewilding of America. University of California Press. ISBN 9780520231412.
- «Fossilworks: Neuryurus». fossilworks.org. Consultado el 24 de enero de 2016.
- «Fossilworks: Parapanochthus». fossilworks.org. Consultado el 24 de enero de 2016.
- Góis, Flávio; Ruiz, Laureano Raúl González; Scillato-Yané, Gustavo Juan; Soibelzon, Esteban (17 de junio de 2015). «A Peculiar New Pampatheriidae (Mammalia: Xenarthra: Cingulata) from the Pleistocene of Argentina and Comments on Pampatheriidae Diversity». PLOS ONE 10 (6): e0128296. ISSN 1932-6203. PMC 4470999. PMID 26083486. doi:10.1371/journal.pone.0128296.
- Elias, Scott; Mock, Cary (25 de marzo de 2013). Encyclopedia of Quaternary Science. Newnes. ISBN 9780444536426.
- «Insularity and Its Effect on Mammal Evolution». ResearchGate (en inglés). Consultado el 12 de marzo de 2017.
- «Paleozoogeography of the Pleistocene mammals from the Aegean». ResearchGate (en inglés). Consultado el 13 de marzo de 2017.
- Ambrosetti, P., (1968). «The Pleistocene dwarf elephants of Spinagallo (Siracusa, south-eastern Sicily)». Geol. Rom. 7, 277-398.
- Falconer, H. (1868). «On the fossil remains of Elephas melitensis. An extinct pygmy species of Elephant and of other Mammalia, from the ossiferous caves of Malta.». Palaeontol. Memoirs and Notes 2, 292-307.
- «Extinct Pygmy Hippopotamus and Early Man in Cyprus (PDF Download Available)». ResearchGate (en inglés). Consultado el 12 de marzo de 2017.
- «Origin and taxonomy of the fossil elephants of the island of Crete (Greece): Problems and perspectives (PDF Download Available)». ResearchGate (en inglés). Consultado el 13 de marzo de 2017.
- «Die entdeckung von Zwergelefanten in der Höhle "Charkadio" auf der Insel Tilos (Dodekanes, Griechenland)». ResearchGate (en inglés). Consultado el 13 de marzo de 2017.
- «Faune mammologiche dell'isole di Kos e di Samos». ResearchGate (en inglés). Consultado el 13 de marzo de 2017.
- Deperet, C., Mayet, L. (1923). «Les elephants pliocenes. II. Monographie des elephants pliocenes de l’Europe et de l’Afrique du Nord.». Ann. Univ. Lyon 1, 91-224.
- «Uber einen Pleistozanen Zwerge-lefanten von der Insel Naxos (Kykladen)». ResearchGate (en inglés). Consultado el 13 de marzo de 2017.
- «The importance of fossil mammals in reconstructing paleogeography with special reference to the Pleistocene Aegean Archipelago». ResearchGate (en inglés). Consultado el 13 de marzo de 2017.
- «Die pleistozänen Säugetierfaunen der ostmediterranen Inseln». ResearchGate (en inglés). Consultado el 13 de marzo de 2017.
- «Erstmalige Funde von Zwergelefanten auf dem Insel Rhodos». ResearchGate (en inglés). Consultado el 13 de marzo de 2017.
- «Elephas tiliensis n. sp. from Tilos island (Dodecanese, Greece)». ResearchGate (en inglés). Consultado el 17 de marzo de 2017.
- Sondaar, P. Y., Sanges, M., Kotsak.is, T. & de Boer, P. L. (1986). «The Plecistocene deer hunter of Sardinia». Geobios 19 (1 ), 17-25.
- «Late Pleistocene deer fossils from Corbeddu Cave : implications for human colonization of the island of Sardinia / Gerard Klein Hofmeijer, in association with Ministero per i beni culturali e ambientali, Soprintendenza archeologica per le province di Sassari e Nuoro, Italia. - Version details». Trove (en inglés). Consultado el 16 de marzo de 2017.
- «Pleistocene humans in the island environment of Sardinia | Sardoa d-library». www.sardoa.eu. Archivado desde el original el 17 de marzo de 2017. Consultado el 16 de marzo de 2017.
- Delussu, Fabrizio. Lo stato attuale degli studi sulle faune oloceniche della Sardegna centro-settentrionale (en inglés). Consultado el 16 de marzo de 2017.
- Delussu, F. (1996). «I resli faunislici. ln (M. Milanese, Ed.) ll vilaggio medievale di Geridu (Sorso, SS).». Sassari: Universita degli studi, pp. 530-548.
- «Cypriot Hippo Hunters No Myth». ResearchGate (en inglés). Consultado el 12 de marzo de 2017.
- Guilaine, J., Coularou, J., Briois, F. & Carrere, I. (1995). «L'etablissement neolilhiquc de Shillourokambos (Parekklisha, Chypre).». Departmem of Antiquities Cyprus.
- Boekschoten, G. J. & Sondaar, P. Y. (1972). «On the fossil mammalia of Cyprus.». Proceedings of the Kononklijke Nederlandse Akad. van Wetenschappen.
- «Search for images at Natural History Museum Picture Library». piclib.nhm.ac.uk. Archivado desde el original el 19 de abril de 2016. Consultado el 20 de abril de 2016.
- «Timing of Hot Spot--Related Volcanism and the Breakup of Madagascar and India (PDF Download Available)». ResearchGate (en inglés). Consultado el 17 de marzo de 2017.
- «Biogeographic evolution of Madagascar's micro-endemic biota (PDF Download Available)». ResearchGate (en inglés). Consultado el 17 de marzo de 2017.
- «On the Origins and Admixture of Malagasy: New Evidence from High-Resolution Analyses of Paternal and Maternal Lineages (PDF Download Available)». ResearchGate (en inglés). Consultado el 17 de marzo de 2017.
- Laurie R. Godfrey M. T Irwin Gary T. Schwartz W. L Jungers (2008). «Ghosts and Orphans: Madagascar's Vanishing Ecosystem». Elwyn Simons: A Search for Origins.
- Burney, David A. (1 de enero de 1999). MacPhee, Ross D. E., ed. Extinctions in Near Time. Advances in Vertebrate Paleobiology (en inglés). Springer US. pp. 145-164. ISBN 9781441933157. doi:10.1007/978-1-4757-5202-1_7#page-1. Consultado el 17 de marzo de 2017.
- «Extinct New Zealand megafauna were not in decline before human colonization (PDF Download Available)». ResearchGate (en inglés). Consultado el 18 de marzo de 2017.
- «Mechanics of overkill in the extinction of New Zealand moas». ResearchGate (en inglés). Consultado el 18 de marzo de 2017.
- «Rapid Extinction of the Moas (Aves: Dinornithiformes): Model, Test, and Implications». ResearchGate (en inglés). Consultado el 18 de marzo de 2017.
- M. Buncea,1,2, T. H. Worthyb,c,1, M. J. Phillipsd, R. N. Holdawaye, E. Willerslevf , J. Hailef,g, B. Shapirog,h, R. P. Scofieldi , A. Drummondj , P. J. J. Kampk, and A. Cooper (2009). «The evolutionary history of the extinct ratite moa and New Zealand Neogene paleogeography». University of California, Santa Barbara, CA.
- «Molecular and morphological analyses of avian eggshell excavated from a late thirteenth century earth oven (PDF Download Available)». ResearchGate (en inglés). Consultado el 18 de marzo de 2017.
- White, Arthur W.; Worthy, Trevor H.; Hawkins, Stuart; Bedford, Stuart; Spriggs, Matthew (31 de agosto de 2010). «Megafaunal meiolaniid horned turtles survived until early human settlement in Vanuatu, Southwest Pacific». Proceedings of the National Academy of Sciences (en inglés) 107 (35): 15512-15516. ISSN 0027-8424. PMC 2932593. PMID 20713711. doi:10.1073/pnas.1005780107. Consultado el 24 de marzo de 2017.
- Keim, Brandon. «Extinct, King Koopa-Style Giant Turtle Found on Pacific Island». WIRED (en inglés estadounidense). Consultado el 24 de marzo de 2017.
- Atholl, Anderson,; Christophe, Sand,; Fiona, Petchey,; H., Worthy, Trevor (1 de enero de 2010). Faunal extinction and human habitation in New Caledonia: Initial results and implications of new research at the Pindai Caves (en inglés) 1. Consultado el 25 de marzo de 2017.
- «Historical Records of Indian Ocean Giant Tortoise Populations». ResearchGate (en inglés). Consultado el 17 de marzo de 2017.
- «Conservation in Mauritius and Rodrigues: Challenges and Achievements from Two Ecologically Devastated Oceanic Islands (PDF Download Available)». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- «Ancient mitochondrial DNA and morphology elucidate an extinct island radiation of Indian Ocean giant tortoises (Cylindraspis) (PDF Download Available)». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- «Galapagos Tortoise Lonesome George May Have Company». Live Science. Consultado el 25 de marzo de 2017.
- Steadman, David W. (1986). «Holocene vertebrate fossils from Isla Floreana, Galápagos». Consultado el 25 de marzo de 2017.
- Poulakakis, Nikos; Glaberman, Scott; Russello, Michael; Beheregaray, Luciano B.; Ciofi, Claudio; Powell, Jeffrey R.; Caccone, Adalgisa (7 de octubre de 2008). «Historical DNA analysis reveals living descendants of an extinct species of Galápagos tortoise». Proceedings of the National Academy of Sciences (en inglés) 105 (40): 15464-15469. ISSN 0027-8424. PMC 2563078. PMID 18809928. doi:10.1073/pnas.0805340105. Consultado el 25 de marzo de 2017.
- «Chelonoidis phantastica». The Reptile Database. Consultado el 25 de marzo de 2017.
- California Academy of Sciences (1 de enero de 1907). Expedition of the California academy of sciences to the Galapagos islands, 1905-1906. San Francisco, The Academy. Consultado el 25 de marzo de 2017.
- Worthy, Trevor. «Pregill G.K. & T.H. Worthy, 2003. A new iguanid lizard (Squamata, Iguanidae) from the late Quaternary of Fiji, Southwest Pacific. Herpetologica 59: 57-67.». Herpetologica (en inglés). Consultado el 24 de marzo de 2017.
- «South Pacific Iguanas: Human Impacts and a New Species». ResearchGate (en inglés). Consultado el 24 de marzo de 2017.
- «The Recently Extinct Plants and Animals Database cubit: The Recently Extinct Plants and Animals Database Extinct Reptiles: Mekosuchus inexpectatus». cubits.org. Archivado desde el original el 4 de marzo de 2016. Consultado el 24 de marzo de 2017.
- «New Extinct Mekosuchine Crocodile from Vanuatu, South Pacific (PDF Download Available)». ResearchGate (en inglés). Consultado el 24 de marzo de 2017.
- «Faunal extinction and human habitation in New Caledonia: Initial results and implications of new research at the Pindai Caves (PDF Download Available)». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- Worthy, Trevor. Worthy, T.H., 2000. The fossil megapodes (Aves: Megapodiidae) of Fiji with descriptions of a new genus and two new species.Journal of the Royal Society of New Zealand 30(4): 337-364. (en inglés). Consultado el 25 de marzo de 2017.
- «A giant flightless pigeon gen. et sp. Nov. and a new species of Ducula (Aves: Columbidae), from Quaternary deposits in Fiji (PDF Download Available)». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- «A new species of extinct imperial pigeon (Ducula: Columbidae) from Henderson Island, Pitcairn Group (PDF Download Available)». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- Steadman, David W. (1992). «New species of Gallicolumba and Macropygia (Aves: Columbidae) from archeological sites in Polynesia». Science series, Natural History Museum of Los Angeles County. 36: 329–348.
- «A new species of Gallicolumba: Columbidae from Henderson Island, Pitcairn Group (PDF Download Available)». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- «Human Dispersal and Late Quaternary Megafaunal Extinctions: the Role of the Americas in the Global Puzzle (PDF Download Available)». ResearchGate (en inglés). Consultado el 21 de febrero de 2017.
- Wenban-Smith, F. F.; Allen, P.; Bates, M. R.; Parfitt, S. A.; Preece, R. C.; Stewart, J. R.; Turner, C.; Whittaker, J. E. (1 de julio de 2006). «The Clactonian elephant butchery site at Southfleet Road, Ebbsfleet, UK». Journal of Quaternary Science (en inglés) 21 (5): 471-483. ISSN 1099-1417. doi:10.1002/jqs.1033. Consultado el 16 de marzo de 2017.
- «Historic extinctions: a rosetta stone for understanding prehistoric extinctions.». ResearchGate (en inglés). Consultado el 11 de marzo de 2017.
- JOSÉ MARÍA FERNÁNDEZ-PALACIOS (2004). «INTRODUCCIÓN A LAS ISLAS». http://jmferpal.webs.ull.es. ECOLOGÍA INSULAR / ISLAND ECOLOGY. ASOCIACIÓN ESPAÑOLA DE ECOLOGÍA TERRESTRE (AEET). Consultado el 21 de febrero de 2017.
- «Leopard seal predation rates at penguin colonies of different size (PDF Download Available)». ResearchGate (en inglés). Consultado el 11 de marzo de 2017.
- «Mammoths: Giants of the Ice Age». ResearchGate (en inglés). Consultado el 24 de febrero de 2017.
- Doughty, C. E.; Wolf, A.; Malhi, Y. (11 de agosto de 2013). «The legacy of the Pleistocene megafauna extinctions on nutrient availability in Amazonia». Nature Geoscience. doi:10.1038/ngeo1895.
- «Pleistocene Extinctions: The Pivotal Role of Megaherbivores». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- «Rates, patterns, and processes of landscape transformation and extinction in Madagascar». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- Marshall, M. (11 de agosto de 2013). «Ecosystems still feel the pain of ancient extinctions». New Scientist. Consultado el 12 de agosto de 2013.
- «Combining paleo-data and modern exclosure experiments to assess the impact of megafauna extinctions on woody vegetation (PDF Download Available)». ResearchGate (en inglés). Consultado el 26 de marzo de 2017.
- «Consumer control by megafauna and fire». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- «Wolves, elk, and aspen in the winter range of Jasper National Park, Canada (PDF Download Available)». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- «Serengeti National Park, Tanzania (PDF Download Available)». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- «Interference competition between gray wolves and coyotes in Yellowstone National Park (PDF Download Available)». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- «Sergey Zimov: lessons from a Pleistocene rewilding in Siberia». Oxford Megafauna Meeting. Consultado el 20 de febrero de 2017.
- «The Economic Value of Elephants (PDF Download Available)». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- «Effects of elephants on ecosystems and biodiversity (PDF Download Available)». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- «Tree-grass dynamics in a Prosopis-thornscrub savanna parkland: Reconstructing the past and predicting the future (PDF Download Available)». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- «Realistic parameter assessment for a well known elephant–tree ecosystem model reveals that limit cycles are unlikely (PDF Download Available)». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- «Sporormiella fungal spores, a palynological means of detecting herbivore density». ResearchGate. Consultado el 25 de marzo de 2017.
- «The impact of the megafauna extinctions on savanna woody cover in South America (PDF Download Available)». ResearchGate. Consultado el 26 de marzo de 2017.
- «High-Resolution Global Maps of 21st-Century Forest Cover Change (PDF Download Available)». ResearchGate (en inglés). Consultado el 26 de marzo de 2017.
- «Body mass of Late Quaternary mammals (PDF Download Available)». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- Owen-Smith, N. (2013). «Contrasts in the large herbivore faunas of the southern continents in the late Pleistocene and the ecological implications for human origins». J Biogeogr 40: 1215-1224.
- «What controls the distribution of tropical forest and savanna? Ecol Lett (PDF Download Available)». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- «Review of feeding ecology data of Late Pleistocene mammalian herbivores from South America and discussions on niche differentiation (PDF Download Available)». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- «Parks of the Pleistocene: recreating the Cerrado and the Pantanal with megafauna (PDF Download Available)». ResearchGate (en inglés). Consultado el 26 de marzo de 2017.
- «Pleistocene Megafaunal Collapse, Novel Plant Communities, and Enhanced Fire Regimes in North America (PDF Download Available)». ResearchGate. Consultado el 26 de marzo de 2017.
- «Late-Quaternary vegetation dynamics in North America: scaling from taxa to biomes». ResearchGate (en inglés). Consultado el 26 de marzo de 2017.
- JOHN W. WILLIAMS, BRYAN N. SHUMAN, AND THOMPSON WEBB III (Ecology, 82(12), 2001, pp. 3346–3362). «DISSIMILARITY ANALYSES OF LATE-QUATERNARY VEGETATION AND CLIMATE IN EASTERN NORTH AMERICA». 2001.
- Williams, John W.; Jackson, Stephen T. (1 de noviembre de 2007). «Novel climates, no-analog communities, and ecological surprises». Frontiers in Ecology and the Environment (en inglés) 5 (9): 475-482. ISSN 1540-9309. doi:10.1890/070037. Consultado el 26 de marzo de 2017.
- «A new Greenland ice core chronology for the Last Glacial termination (PDF Download Available)». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- Summerhayes, Glenn R.; Leavesley, Matthew; Fairbairn, Andrew; Mandui, Herman; Field, Judith; Ford, Anne; Fullagar, Richard (1 de octubre de 2010). «Human adaptation and plant use in highland New Guinea 49,000 to 44,000 years ago». Science (New York, N.Y.) 330 (6000): 78-81. ISSN 1095-9203. PMID 20929808. doi:10.1126/science.1193130. Consultado el 26 de marzo de 2017.
- Bowler, James M.; Johnston, Harvey; Olley, Jon M.; Prescott, John R.; Roberts, Richard G.; Shawcross, Wilfred; Spooner, Nigel A. «New ages for human occupation and climatic change at Lake Mungo, Australia». Nature 421 (6925): 837-840. doi:10.1038/nature01383.
- Turney, Chris S. M.; Flannery, Timothy F.; Roberts, Richard G.; Reid, Craig; Fifield, L. Keith; Higham, Tom F. G.; Jacobs, Zenobia; Kemp, Noel et al. (26 de agosto de 2008). «Late-surviving megafauna in Tasmania, Australia, implicate human involvement in their extinction». Proceedings of the National Academy of Sciences (en inglés) 105 (34): 12150-12153. ISSN 0027-8424. PMC 2527880. PMID 18719103. doi:10.1073/pnas.0801360105. Consultado el 26 de marzo de 2017.
- «Pleistocene faunal loss: Implications of the aftershock for Australia's past and future». ResearchGate (en inglés). Consultado el 26 de marzo de 2017.
- «Even carbon number predominance of plant wax n-alkanes: A correction». ResearchGate (en inglés). Consultado el 26 de marzo de 2017.
- «Preindustrial Human Impacts on the Global/Regional Environment». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- O’Connor, M.; Wolf, A.; Doughty, C. E.; Malhi, Y. (9 de agosto de 2013). «Lateral Diffusion of Nutrients by Mammalian Herbivores in Terrestrial Ecosystems». PLoS ONE 8 (8): e71352. doi:10.1371/journal.pone.0071352.
- Daniel H. Janzen and Paul S. Martin (1982). «Neotropical Anachronisms: The Fruits the Gomphotheres Ate». Science 215(4528):19-27.
- «Reconstructing past ecological networks: The reconfiguration of seed-dispersal interactions after megafaunal extinction (PDF Download Available)». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- «Megagardeners of the forest - the role of elephants in seed dispersal (PDF Download Available)». ResearchGate (en inglés). Consultado el 29 de marzo de 2017.
- «Forest Elephants: Tree Planters of the Congo (PDF Download Available)». ResearchGate (en inglés). Consultado el 29 de marzo de 2017.
- «Asian Tapirs Are No Elephants When It Comes To Seed Dispersal (PDF Download Available)». ResearchGate (en inglés). Consultado el 29 de marzo de 2017.
- «Gourds and squashes (Cucurbita spp.) adapted to megafaunal extinction and ecological anachronism through domestication (PDF Download Available)». ResearchGate (en inglés). Consultado el 29 de marzo de 2017.
- «Megafauna extinction, tree species range reduction, and carbon storage in Amazonian forests (PDF Download Available)». ResearchGate (en inglés). Consultado el 22 de febrero de 2017.
- Walter, K. M.; Zimov, S. A.; Chanton, J. P.; Verbyla, D.; Chapin, F. S. (7 de septiembre de 2006). «Methane bubbling from Siberian thaw lakes as a positive feedback to climate warming». Nature (en inglés) 443 (7107): 71-75. ISSN 0028-0836. doi:10.1038/nature05040. Consultado el 3 de abril de 2017.
- «Estimation of the methane emission factor for the Italian Mediterranean buffalo (PDF Download Available)». ResearchGate (en inglés). Consultado el 28 de marzo de 2017.
- Smith, F. A.; Elliot, S. M.; Lyons, S. K. (23 de mayo de 2010). «Methane emissions from extinct megafauna». Nature Geoscience (Nature Publishing Group) 3 (6): 374-375. doi:10.1038/ngeo877. Archivado desde el original el 30 de marzo de 2011. Consultado el 26 de febrero de 2011.
- «Methane emissions from bison—An historic herd estimate for the North American Great Plains». ResearchGate (en inglés). Consultado el 28 de marzo de 2017.
- «Late Pleistocene Megafaunal Extinctions». ResearchGate (en inglés). Consultado el 21 de febrero de 2017.
- Kelliher, F. M.; Clark, H. (15 de marzo de 2010). «Methane emissions from bison—An historic herd estimate for the North American Great Plains». Agricultural and Forest Meteorology 150 (3): 473-577. doi:10.1016/j.agrformet.2009.11.019.
- Zalasiewicz, Jan; Waters, Colin N.; Williams, Mark; Barnosky, Anthony D.; Cearreta, Alejandro; Crutzen, Paul; Ellis, Erle; Ellis, Michael A. et al. (5 de octubre de 2015). «When did the Anthropocene begin? A mid-twentieth century boundary level is stratigraphically optimal». Quaternary International. The Quaternary System and its formal subdivision 383: 196-203. doi:10.1016/j.quaint.2014.11.045. Consultado el 26 de marzo de 2017.
- «Marine defaunation: Animal loss in the global ocean (PDF Download Available)». ResearchGate (en inglés). Consultado el 26 de marzo de 2017.
- «Effects of mammalian herbivore declines on plant communities: Observations and experiments in an African savanna (PDF Download Available)». ResearchGate (en inglés). Consultado el 26 de marzo de 2017.
- «Defaunation in the Anthropocene (PDF Download Available)». ResearchGate (en inglés). Consultado el 26 de marzo de 2017.
- «Collapse of the world's largest herbivores (PDF Download Available)». ResearchGate (en inglés). Consultado el 26 de marzo de 2017.
- «Rewilding: Science, Practice, and Politics». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- «Rewilding with large herbivores: The importance of grazing refuges for sapling establishment and wood-pasture formation (PDF Download Available)». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- «Release of Arnold's giant tortoises Dipsochelys arnoldi on Silhouette island, Seychelles (PDF Download Available)». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- «Paper: Sterilisation of hybrid Galapagos tortoises (Geochelone nigra) for Island restoration. Part 1: Endoscopic oophorectomy of females under ketamine-medetomidine anaesthesia (PDF Download Available)». ResearchGate (en inglés). Consultado el 25 de marzo de 2017.
- «Rethinking rewilding». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- Sandom C, Donlan CJ, Svenning J-C, Hansen D. (2013). «Rewilding.». Key Topics in Conservation Biology 2:430-51: John Wiley & Sons. Number of 430-51 pp.
- Soule M, Noss R. (1998). «Rewilding and biodiversity: complementary goals for continental conservation.». Wild Earth Fall 22.
- Soule M, Terborgh J. (1999). «Continental Conservation: Scientific Foundations of Regional Reserve Networks.». Island Press.
- «Trophic cascades in Yellowstone: The first 15 years after wolf reintroduction». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- «Is science in danger of sanctifying the wolf Biol». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- «Ecological history and latent conservation potential: large and giant tortoises as a model for taxon substitution (PDF Download Available)». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- «Rewilding North America (PDF Download Available)». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- «Demographic Outcomes and Ecosystem Implications of Giant Tortoise Reintroduction to Espanola Island, Galapagos (PDF Download Available)». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- «Assessing the Potential to Restore Historic Grazing Ecosystems with Tortoise Ecological Replacements (PDF Download Available)». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- «Rewilding is the new Pandora's box in conservation (PDF Download Available)». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- «Vertebrate tracks and trackways from the pleistocene eolianites of SW Portugal (PDF Download Available)». ResearchGate (en inglés). Consultado el 28 de marzo de 2017.
- «Vertebrate tracks of the Late Pleistocene eolianites from SW Alentejo, Portugal». ResearchGate (en inglés). Consultado el 28 de marzo de 2017.
- «Vertebrate tracksites from the Mid-Late Pleistocene eolianites of Portugal: The first record of elephant tracks in Europe (PDF Download Available)». ResearchGate (en inglés). Consultado el 28 de marzo de 2017.
- «Applying Landscape Ecology in Biological Conservation». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- «Agricultural Abandonment in Mountain Areas of Europe: Environmental Consequences and Policy Response (PDF Download Available)». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- «Rewilding Abandoned Landscapes in Europe (PDF Download Available)». ResearchGate (en inglés). Consultado el 28 de marzo de 2017.
- «Grazing Ecology and Forest History (PDF Download Available)». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- «The role of large grazing animals in nature conservation - A Dutch perspective». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- «Really wild? Naturalistic grazing in modern landscapes». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- «A review of natural vegetation openness in Northwestern Europe». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- «Can the pre-Neolithic provide suitable models for rewilding the landscape in Britain? (PDF Download Available)». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- «How open were European primeval forests? Hypothesis testing using palaeoecological data (PDF Download Available)». ResearchGate. Consultado el 27 de marzo de 2017.
- «High herbivore density associated with vegetation diversity in interglacial ecosystems (PDF Download Available)». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- «The potential role of large herbivores in nature conservation and extensive land use in Europe.». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- Goderie R, Helmer W, Kerkdijk-Otten H, Widstrand S. (2013). The Aurochs: Born to be Wild. The Netherlands: Roodbont.
- «Reintroducing resurrected species: Selecting DeExtinction candidates». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- Europe R. (2012). «Rewilding Europe: Making Europe a Wilder Place.». Nijmegen: Rewilding Europe.
- «Rewilding Europe: A New Strategy for an Old Continent (PDF Download Available)». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- «Rewilding Europe as a new agent of change? Exploring the governance of an experimental discourse and practice in European nature conservation (PDF Download Available)». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- «Ecosystem Services: The Opportunities of Rewilding in Europe (PDF Download Available)». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- JORGE ESTÉVEZ ESCALERA (1978). «Primer hallazgo del buey almizclado (Ovibos moschatus, Zimmermann) en el pleistoceno peninsular». ACTA GEOLÓGICA HISPÁNICA t. XIII (1978), n.O 2, págs. 59-60.
- ICMO2 (2010). «Natural processes, animal welfare, moral aspects and management of the Oostvaardersplassen». Report of the second International Commission on Management of the Oostvaardersplassen (ICMO2). The Hague/Wageningen, Netherlands. Wing rapport 039.
- ICMO (2006). «Reconciling Nature and Human Interests: Advice of the International Committee on the Management of large herbivores in the Oostvaardersplassen (ICMO).». The Hague: Wageningen.
- van den Belt, Henk. «Networking nature, or serengeti behind the dikes». History and Technology (en inglés) 20 (3): 311-333. doi:10.1080/0734151042000287023. Consultado el 28 de marzo de 2017.
- Zimov, Sergey A. (6 de mayo de 2005). «Pleistocene Park: Return of the Mammoth's Ecosystem». Science (en inglés) 308 (5723): 796-798. ISSN 0036-8075. PMID 15879196. doi:10.1126/science.1113442. Consultado el 28 de marzo de 2017.
- Zimov, Sergey A.; Zimov, N. S.; Iii, F. S. Chapin (1 de enero de 2012). Louys, Julien, ed. Paleontology in Ecology and Conservation. Springer Earth System Sciences (en inglés). Springer Berlin Heidelberg. pp. 193-225. ISBN 9783642250378. doi:10.1007/978-3-642-25038-5_10#page-1. Consultado el 28 de marzo de 2017.
- «Pleistocene Park crowdfunding (PDF Download Available)». ResearchGate (en inglés). Consultado el 28 de marzo de 2017.
- «Mammoth steppe: A high-productivity phenomenon (PDF Download Available)». ResearchGate (en inglés). Consultado el 28 de marzo de 2017.
- «J. Craig Venter Life at the Speed of Light: From the Double Helix to the Dawn of Digital Life. New York: Viking (2013) 187 pp. G. Church and E. Regis Regenesis: How Synthetic Biology Will Reinvent Nature and Ourselves. New York: Basic Books (2012) 284 pp.». ResearchGate (en inglés). Consultado el 27 de marzo de 2017.
- Mark, Jason. «De-extinction won’t make us better conservationists!». Salon. Consultado el 28 de marzo de 2017.
- «Opinion: The Case Against Species Revival». 12 de marzo de 2017. Consultado el 28 de marzo de 2017.
- Editors, The. «Do Not Reanimate». Scientific American (en inglés) 308 (6): 12-12. doi:10.1038/scientificamerican0613-12. Consultado el 28 de marzo de 2017.
- «Bringing Them Back to Life». 1 de abril de 2013. Consultado el 28 de marzo de 2017.
- Biello, David. «Will we kill off today's animals if we revive extinct ones?». Nature (en inglés). doi:10.1038/nature.2013.12645. Consultado el 28 de marzo de 2017.
- Miller, Webb; Drautz, Daniela I.; Ratan, Aakrosh; Pusey, Barbara; Qi, Ji; Lesk, Arthur M.; Tomsho, Lynn P.; Packard, Michael D. et al. (20 de noviembre de 2008). «Sequencing the nuclear genome of the extinct woolly mammoth». Nature (en inglés) 456 (7220): 387-390. ISSN 0028-0836. doi:10.1038/nature07446. Consultado el 29 de marzo de 2017.
- Campbell, Kevin L.; Roberts, Jason E. E.; Watson, Laura N.; Stetefeld, Jörg; Sloan, Angela M.; Signore, Anthony V.; Howatt, Jesse W.; Tame, Jeremy R. H. et al. (1 de junio de 2010). «Substitutions in woolly mammoth hemoglobin confer biochemical properties adaptive for cold tolerance». Nature Genetics (en inglés) 42 (6): 536-540. ISSN 1061-4036. doi:10.1038/ng.574. Consultado el 29 de marzo de 2017.
- «Scientists Make Progress on Building a More Mammoth-Like Elephant - NBC News». NBC News (en inglés). Consultado el 28 de marzo de 2017.
- «Scientists Want to Bring Back Woolly Mammoths — But Why?». NBC News (en inglés). Consultado el 28 de marzo de 2017.
- «'Initial stage' reached on dream of cloning woolly mammoth - expert». siberiantimes.com (en inglés). Consultado el 28 de marzo de 2017.
- «Can scientists bring mammoths back to life?». Washington Post. Consultado el 29 de marzo de 2017.
- «Russia: New laboratory to study mammoth cloning». BBC News (en inglés británico). 1 de septiembre de 2015. Consultado el 28 de marzo de 2017.
- Can scientists clone extinct Ice Age cave lion? (en inglés británico), 4 de marzo de 2016, consultado el 28 de marzo de 2017.
- «The remains of an 8,000 year old lunch: an extinct steppe bison's tail». siberiantimes.com (en inglés). Consultado el 28 de marzo de 2017.
- «Century After Extinction, Passenger Pigeons Remain Iconic—And Scientists Hope to Bring Them Back». 30 de agosto de 2017. Consultado el 28 de marzo de 2017.
- «Pictures: Extinct Species That Could Be Brought Back». 5 de marzo de 2013. Consultado el 28 de marzo de 2017.
- News, A. B. C. (7 de enero de 2006). «Extinct Tasmanian Tiger Could Be Cloned». ABC News. Consultado el 28 de marzo de 2017.
- Zaragoza, Víctor Millán. «¿Y si Aragón hubiese logrado clonar al bucardo?». heraldo.es. Consultado el 28 de marzo de 2017.
- «Sasha, the world's only baby woolly rhino, is 34,000 years old, say scientists». siberiantimes.com (en inglés). Consultado el 28 de marzo de 2017.
| Control de autoridades |
|
|---|
 Datos: Q1759232
Datos: Q1759232