Sistema inmunitario
El sistema inmunitario o sistema inmunológico es el conjunto de elementos y procesos biológicos en el interior de un organismo que le permite mantener la homeostasis o equilibrio interno frente a agresiones externas, ya sean de naturaleza biológica (agentes patógenos) o físicoquímicas (como contaminantes o radiaciones) e internas (por ejemplo, células cancerosas).[1][2][3] Reconoce lo dañino y reacciona frente a ello (ya sea una agresión externa o interna).[1]
| Sistema inmunitario | ||
|---|---|---|
 En la imagen, neutrófilos (en color amarillo) fagocitando bacterias del carbunco (en color naranja). Imagen obtenida mediante un microscopio electrónico de barrido. La línea blanca en la parte inferior izquierda equivale a 5 micrómetros. | ||
| Estudiado (a) por | inmunología | |
| Información fisiológica | ||
| Función | Protección de un organismo ante agentes externos. | |
| Estructuras principales | ||
| Leucocitos o glóbulos blancos | ||
|
| ||
El sistema inmunitario lo componen moléculas solubles (como las proteínas del sistema complemento, los anticuerpos, la histamina, etcétera) en diferentes fluidos (sangre y linfa, entre otros) y células localizadas en diferentes tejidos y órganos, principalmente: médula ósea, timo, bazo, ganglios linfáticos y MALT o tejido linfoide asociado a las mucosas. En la médula ósea se generan distintos tipos de leucocitos o glóbulos blancos, que son células especializadas en la función inmune: neutrófilos, linfocitos, eosinófilos, basófilos, mastocitos, monocitos, células dendríticas y macrófagos; todas ellas se movilizan a través de la sangre y el sistema linfático hacia los distintos órganos.[4][5][6][7][8][9]
Hay diferentes niveles en la respuesta inmunitaria. Algunos elementos de respuesta son invariantes en el tiempo y se agrupan en la denominada respuesta inmunitaria innata (natural o inespecífico) y alternativamente los elementos capaces de memorizar a los microorganismos se organizan en la respuesta inmunitaria adquirida (adaptativo o específico). Los elementos de respuesta innata están presentes prácticamente en todos los seres vivos, incluso los sencillos organismos unicelulares como las bacterias poseen sistemas enzimáticos que los protegen contra infecciones virales. Otros mecanismos inmunitarios básicos se encuentran en eucariontes, plantas, peces, reptiles e insectos, así como en mamíferos. Entre estos mecanismos figuran péptidos antimicrobianos llamados defensinas y citocinas, la fagocitosis que realizan neutrófilos y macrófagos, el sistema del complemento y otros. El sistema inmunitario innato puede detectar en las células una variedad de señales de «peligro» llamadas patrones moleculares asociados a peligro (DAMP, por sus siglas del inglés) o bien la presencia de señales asociadas a agentes patógenos denominadas patrones moleculares asociados a patógenos (PAMP, por sus siglas del inglés), identificando de esta forma una amplia variedad de células dañadas, ya sea por quemaduras, radiación, virus, bacterias, parásitos y muchos otros agentes, distinguiéndolas de las células y tejidos sanos del organismo para funcionar correctamente.[1][4][10][11][12]
La respuesta inmunitaria adquirida permite que los vertebrados, como los humanos, tengan mecanismos de defensa más sofisticados, interconectados con los mecanismos del sistema inmunitario innato en forma dinámica y de más largo plazo. La unidad anatómico-funcional de ese sistema es el linfocito. El sistema inmunitario se adapta con el tiempo para reconocer patógenos específicos de manera más eficaz, generando una memoria inmunitaria. La memoria inmunitaria creada desde una respuesta primaria a un patógeno específico proporciona una respuesta mejorada a encuentros secundarios con ese mismo patógeno específico. Este proceso de inmunidad adquirida es la base de la vacunación.[1][13][14]
Los trastornos en el sistema inmunitario pueden ocasionar muchas enfermedades. La inmunodeficiencia ocurre cuando el sistema inmunitario es menos activo que lo normal,[15] lo que favorece las infecciones recidivantes y con peligro para la vida. La inmunodeficiencia puede ser el resultado de una enfermedad genética, como la inmunodeficiencia combinada grave,[16] o ser producida por fármacos o una infección, como el síndrome de inmunodeficiencia adquirida (sida) que está provocado por el retrovirus VIH.[17] En cambio, las enfermedades autoinmunes son consecuencia de un sistema inmunitario hiperactivo que ataca tejidos normales como si fueran organismos extraños. Entre las enfermedades autoinmunitarias comunes figuran la tiroiditis de Hashimoto, la artritis reumatoide, la diabetes mellitus tipo 1 y el lupus eritematoso. La inmunología cubre el estudio de todos los aspectos del sistema inmunitario que tienen relevancia significativa para la salud humana y las enfermedades. Se espera que la mayor investigación en este campo desempeñará un papel importante en la promoción de la salud y el tratamiento de enfermedades.

Terminología
El vocablo «inmunidad» es un neologismo del siglo XIX derivado del latín in-mūn(itātem) 'sin obligación', cuyo sentido actual se remonta al año 1866, probablemente influido por el término latino de uso militar immunīre 'defender desde dentro'.[18]
Los términos relacionados «inmunitario» (perteneciente o relativo a la inmunidad), «inmunológico» (perteneciente o relativo a la inmunología), «inmunología» (estudio de la inmunidad biológica y sus aplicaciones) e «inmune» (no atacable por ciertas enfermedades; o bien, perteneciente o relativo a las causas, mecanismos o efectos de la inmunidad), son todos vocablos aceptados por la RAE.
Respecto al uso académico, en concordancia con las definiciones de la RAE, es correcto referirse tanto a sistema inmunitario como a sistema inmune, ya que en este último caso la palabra inmune toma la acepción de «perteneciente o relativo a las causas, mecanismos o efectos de la inmunidad» (Nota: en el inglés «immune» se usa tanto para inmune como inmunitario). No obstante, el uso de sistema inmunológico puede observarse en numerosas publicaciones y obras de referencia en el idioma español.[19][21]
Órganos primarios y secundarios
El sistema inmunitario consta de una serie de órganos, tejidos y células ampliamente repartidos por todo el cuerpo. Funcionalmente, los órganos se clasifican en primarios y secundarios. Los primarios son la médula ósea y el timo, que son los que proporcionan el microambiente para la maduración de los linfocitos de la inmunidad adquirida. Los órganos secundarios son los ganglios linfáticos, el bazo, el tejido linfoide asociado a las mucosas (y en parte la propia médula ósea) en donde las células inmunitarias migran con el microorganismo o antígeno en su interior, suministrando el entorno adecuado para que los linfocitos interactúen con él, en el proceso denominado presentación de antígeno.[22]
Líneas inmunitarias de defensa
El sistema inmunitario protege los organismos de las infecciones con varias líneas de defensa de especificidad creciente. La primera línea de defensa la constituyen las barreras físicas, químicas y biológicas que son la piel y todas las superficies mucosas del organismo (tracto gastrointestinal, respiratorio, genitourinario, así como la conjuntiva ocular), que evitan que patógenos como bacterias y virus entren en el organismo. Si un patógeno penetra estas barreras, la inmunidad innata ofrece una respuesta inmediata, pero no específica. Los elementos innatos de respuesta existen en todas las plantas y animales.[23] Sin embargo, si los agentes patógenos evaden la respuesta innata, los vertebrados poseen un tercer nivel de defensa, que es la respuesta inmunitaria adquirida. Aquí el sistema inmunitario adapta su respuesta durante la infección para mejorar el reconocimiento del agente patógeno. La información sobre esta respuesta mejorada se conserva aun después de que el agente patógeno sea eliminado, bajo la forma de memoria inmunitaria, y permite que el sistema inmunitario adaptativo desencadene ataques más rápidos y más fuertes si en el futuro el sistema inmunitario detecta este tipo de patógeno.[24]
Características del sistema inmunitario
| Inmunidad Innata | Inmunidad adaptativa |
| La respuesta no es específica. | Respuesta específica. |
| Se responde a Patrones Moleculares Asociados a Patógenos (PAMPs). | Se responde a antígenos de los patógenos. |
| La exposición conduce a la respuesta máxima inmediata. | Demora entre la exposición y la respuesta máxima. |
| Inmunidad mediada por células y componentes humorales. | Inmunidad mediada por células y componentes humorales. |
| Sin memoria inmunológica. | La exposición conduce a la memoria inmunológica. |
| Presente en casi todas las formas de vida. | Presente solo en vertebrados mandibulados. |
Tanto la inmunidad innata como la adaptativa dependen de la habilidad del sistema inmunitario para distinguir entre las moléculas propias y las que no lo son. En inmunología, las moléculas propias son aquellos componentes de un organismo que el sistema inmunitario distingue de las substancias extrañas.[25] Al contrario, las moléculas que no son parte del organismo, son reconocidas como moléculas extrañas. Un tipo de moléculas extrañas son los llamados antígenos ("anti", del griego Δντι- que significa 'opuesto' o 'con propiedades contrarias' y "geno", de la raíz griega γεν, generar, producir [que genera o crea oposición]), son substancias que se enlazan a receptores inmunitarios específicos y desencadenan una respuesta inmunitaria.[26] [27]
Inmunidad innata
Los microorganismos o toxinas que consigan entrar en un organismo se encontrarán con las células y los mecanismos del sistema inmunitario innato. La respuesta innata suele desencadenarse cuando los microbios son identificados por receptores de reconocimiento de patrones, que reconocen componentes que están presentes en amplios grupos de microorganismos,[28] o cuando las células dañadas, lesionadas o estresadas envían señales de alarma, muchas de las cuales (pero no todas) son reconocidas por los mismos receptores que reconocen los patógenos.[10] Los gérmenes que logren penetrar en un organismo se encontrarán con las células y los mecanismos del sistema inmunitario innato. Las defensas del sistema inmunitario innato no son específicas, lo cual significa que estos sistemas reconocen y responden a los patógenos en una forma genérica.[26] Este sistema no confiere una inmunidad duradera contra el patógeno. El sistema inmunitario innato es el sistema dominante de protección en la gran mayoría de los organismos.[23]
Barreras superficiales y químicas

Varias barreras protegen a los organismos de las infecciones, incluyendo barreras mecánicas, químicas y biológicas. Las cutículas ceruminosas de muchas hojas, el exoesqueleto de los insectos, las cáscaras y membranas de los huevos puestos en el exterior, y la piel son ejemplos de las barreras mecánicas que forman la primera línea defensiva contra las infecciones.[26] Sin embargo, como los organismos no pueden aislarse completamente de su medio, otros sistemas participan en la protección de las aberturas corporales, como los pulmones, intestinos y el aparato genitourinario. Los pulmones, la tos y los estornudos expulsan mecánicamente los patógenos y otros irritantes de las vías respiratorias. La acción limpiadora de las lágrimas y la orina también expulsa patógenos mecánicamente, mientras que las mucosidades secretadas por los aparatos respiratorio y gastrointestinal sirven para atrapar y enganchar a los microorganismos.[29]
Las barreras químicas también protegen contra infecciones. La piel y el tracto respiratorio secretan péptidos antimicrobianos tales como las defensinas-β.[30] Enzimas tales como la lisozima y la fosfolipasa A en la saliva, las lágrimas y la leche materna también son agentes antibacterianos.[31][32] Las secreciones de la vagina sirven como barreras químicas en la menarquia, cuando se vuelven ligeramente ácidas, mientras que el semen contiene defensinas y zinc para matar patógenos.[33][34] En el estómago, el ácido gástrico y las peptidasas actúan como poderosas defensas químicas frente a patógenos ingeridos.
Dentro de los tractos genitourinario y gastrointestinal, la microbiota comensal sirve como barrera biológica porque compite con las bacterias patógenas por alimento y espacio, y en algunos casos modificando las condiciones del medio, como el pH o el contenido de hierro disponible.[35] Esto reduce la probabilidad de que la población de patógenos alcance el número suficiente de individuos como para causar enfermedades. Sin embargo, dado que la mayoría de los antibióticos no discriminan entre bacterias patógenas y la flora normal, los antibióticos orales pueden a veces producir un crecimiento excesivo de hongos (los hongos no son afectados por la mayoría de los antibióticos) y originar procesos como la candidiasis vaginal (provocada por una levadura).[36] La reintroducción de microorganismos probióticos, como el lactobacillus, encontrado en el yogur, contribuyen a restaurar un equilibrio saludable de las poblaciones microbianas en las infecciones intestinales en los niños, y también hay datos preliminares alentadores en estudios sobre gastroenteritis bacteriana, enfermedades inflamatorias intestinales, infecciones urinarias e infecciones postquirúrgicas.[37][38]
Fiebre
La fiebre, definida como una elevación de la temperatura corporal superior a los 37,7 °C, es, en realidad, una respuesta de protección ante la infección y la lesión,[39] considerada como una estimulación del sistema inmunitario del organismo.[40] La fiebre es provocada por un tipo de monocitos conocidos como pirógenos[41] —siendo sustancias naturales que producen la fiebre—, obligando al cuerpo a que produzca los suyos propios como un modo de defensa ante cualquier infección posible.[42] Sin embargo, las infecciones no son la única causa de la fiebre, a menudo, puede no ser una respuesta inmunológica.[43]
Por lo general, la fiebre tiene una causa obvia como una infección provocada por algún virus, bacteria, hongo o protista, algún tipo de cáncer, una reacción alérgica, trastornos hormonales, ejercicio excesivo, enfermedades autoinmunes, lesión del hipotálamo (glándula endocrina encargada de regular la temperatura del cuerpo; es como un termostato[44]) o por la excesiva exposición al sol. La fiebre, debido a sus potenciales efectos beneficiosos, se discute si debe ser tratado de forma rutinaria.[45][46] La fiebre beneficia al sistema inmunológico para combatir de forma más eficiente a los «invasores»:[47] aumentando y mejorando la movilidad y la fagocitosis de los leucocitos, bajando los niveles de endotoxina, incrementando la proliferación de las células T y mejorando la actividad del interferón.[48][49] La fiebre puede seguir un cuadro en el que alcanza una temperatura máxima diaria y luego regresa a su nivel normal. De igual forma, la fiebre puede ser remitente, es decir, que la temperatura varía, pero no vuelve a la normalidad.
Inflamación
La inflamación es una de las primeras respuestas del sistema inmunitario a una infección.[50] Los síntomas de la inflamación son el enrojecimiento, hinchazón y calor. El enrojecimiento es causado por el incremento del flujo de sangre en el tejido afectado, la hinchazón se debe a la acumulación de células del sistema inmune, las cuales a su vez liberan diferentes sustancias para atacar al patógeno y para continuar reclutando otros leucocitos. Y el calor se genera por el proceso metabólico de las células que están en acción.[51]
La inflamación es producida por eicosanoides y citocinas, que son liberadas por células heridas o infectadas. Los eicosanoides incluyen prostaglandinas que producen fiebre y dilatación de los vasos sanguíneos asociados con la inflamación, y leucotrienos que atraen ciertos leucocitos.[52][53] Las citocinas incluyen interleucinas que son responsables de la comunicación entre los leucocitos; quimiocinas que promueven la quimiotaxis; y los interferones que tienen efectos anti-virales como la supresión de la síntesis de proteínas en la célula huésped.[54] También pueden liberarse factores de crecimiento y factores citotóxicos. Estas citocinas y otros agentes químicos atraen células inmunitarias al lugar de la infección y promueven la curación del tejido dañado mediante la remoción de los patógenos.[55]
Sistema del complemento
El sistema del complemento es una cascada bioquímica que ataca las superficies de las células extrañas. Contiene más de 20 proteínas diferentes y recibe ese nombre por su capacidad para complementar la destrucción de patógenos iniciada por los anticuerpos. El sistema del complemento es el mayor componente humoral de la respuesta inmunitario innata.[9][56] Muchas especies tienen sistemas de complemento, no solo se presenta en los mamíferos, sino que las plantas, peces y algunos invertebrados también lo poseen.[57]
En los seres humanos, esta respuesta es activada por la unión de proteínas del complemento a carbohidratos de las superficies de los microorganismos o por la unión del complemento a anticuerpos que a su vez se han unido a los microorganismos. Esta señal de reconocimiento produce una respuesta rápida de destrucción.[58] La velocidad de la respuesta es el resultado de la amplificación de la señal que ocurre tras la activación proteolítica secuencial de las moléculas del complemento, que también son proteasas. Tras la unión inicial de proteínas del complemento al microbio, aquellas activan su capacidad proteásica, que a su vez activa a otras proteasas del complemento y así sucesivamente. Esto produce una cascada catalítica que amplifica la señal inicial por medio de una retroalimentación positiva controlada.[59] La cascada origina la producción de péptidos que atraen células inmunitarias, aumentan la permeabilidad vascular y opsonizan (recubren) la superficie del patógeno, marcándolo para su destrucción. Esta deposición del complemento puede también matar células directamente al bloquear su membrana plasmática.[9]
Barreras celulares del sistema innato

Los leucocitos (células blancas de la sangre) actúan como organismos unicelulares independientes y son el segundo brazo del sistema inmunitario innato.[26] Los leucocitos innatos incluyen fagocitos (macrófagos, neutrófilos y células dendríticas), mastocitos, eosinófilos, basófilos y células asesinas naturales. Estas células identifican y eliminan patógenos, bien sea atacando a los más grandes a través del contacto o englobando a otros para así matarlos.[57] Las células innatas también son importantes mediadores en la activación del sistema inmunitario adaptativo.[24]
La fagocitosis es una característica importante de la inmunidad innata celular, llevada a cabo por células llamadas fagocitos, que engloban o comen, patógenos y partículas rodeándolos exteriormente con su membrana hasta hacerlos pasar al interior de su citoplasma. Los fagocitos generalmente patrullan en búsqueda de patógenos, pero pueden ser atraídos a ubicaciones específicas por las citocinas.[26] Al ser englobado por el fagocito, el patógeno resulta envuelto en una vesícula intracelular llamada fagosoma que a continuación se fusiona con otra vesícula llamada lisosoma para formar un fagolisosoma. El patógeno es destruido por la actividad de las enzimas digestivas del lisosoma o a consecuencia del llamado "chorro respiratorio" que libera radicales libres de oxígeno en el fagolisosoma.[60][61] La fagocitosis evolucionó como un medio de adquirir nutrientes, pero este papel se extendió en los fagocitos para incluir el englobamiento de patógenos como mecanismo de defensa.[62] La fagocitosis probablemente representa la forma más antigua de defensa del huésped, pues ha sido identificada en animales vertebrados e invertebrados.[63]
Los neutrófilos y macrófagos son fagocitos que viajan a través del cuerpo en busca de patógenos invasores.[64] Los neutrófilos son encontrados normalmente en la sangre y es el tipo más común de fagocitos, que normalmente representan el 50 o 60 % del total de leucocitos que circulan en el cuerpo.[65] Durante la fase aguda de la inflamación, particularmente en el caso de las infecciones bacterianas, los neutrófilos migran hacia el lugar de la inflamación en un proceso llamado quimiotaxis, y son las primeras células en llegar a la escena de la infección. Los macrófagos son células versátiles que residen dentro de los tejidos y producen una amplia gama de sustancias como enzimas, proteínas del complemento, y factores reguladores como la Interleucina 1.[66] Los macrófagos también actúan como carroñeros, librando al organismo de células muertas y otros residuos, y como "células presentadoras de antígenos" para activar el sistema inmunitario adaptativo.[24]
Las células dendríticas son fagocitos en los tejidos que están en contacto con el ambiente externo; por lo tanto están localizados principalmente en la piel, la nariz, los pulmones, el estómago y los intestinos.[67] Se llaman así por su semejanza con las dendritas neuronales, pues ambas tienen muchas proyecciones espiculares en su superficie, pero las células dendríticas no están relacionadas en modo alguno con el sistema nervioso. Las células dendríticas actúan como enlace entre los sistemas inmunitarios innato y adaptativo, pues presentan antígenos a las células T, uno de los tipos de célula clave del sistema inmunitario adaptativo.[67]
Los mastocitos residen en los tejidos conectivos y en las membranas mucosas, y regulan la respuesta inflamatoria.[68] Se encuentran asociadas muy a menudo con la alergia y la anafilaxia.[65] Los basófilos y los eosinófilos están relacionados con los neutrófilos. Secretan mediadores químicos que están involucrados en la defensa contra parásitos y desempeñan un papel en las reacciones alérgicas, como el asma.[69] Las células asesinas naturales (NK, del inglés Natural Killer) son leucocitos que atacan y destruyen células tumorales, o células que han sido infectadas por virus.[70]
Inmunidad adaptativa o adquirida
El sistema inmunitario adaptativo evolucionó en los vertebrados primitivos y permite una respuesta inmunitaria mayor, así como el establecimiento de la denominada "memoria inmunológica", donde cada patógeno es "recordado" por un antígeno característico y propio de ese patógeno en particular.[71] La respuesta inmunitaria adaptativa es específica de los anticuerpos y requiere el reconocimiento de antígenos que no son propios durante un proceso llamado "presentación de los antígenos". La especificidad del antígeno permite la generación de respuestas que se adaptan a patógenos específicos o a las células infectadas por patógenos. La habilidad de montar estas respuestas específicas se mantiene en el organismo gracias a las células de memoria. Si un patógeno infecta a un organismo más de una vez, estas células de memoria desencadenan una respuesta específica para ese patógeno que han reconocido, con el fin de eliminarlo rápidamente.
Linfocitos
Las células del sistema inmunitario adaptativo son una clase especial de leucocitos, llamados linfocitos. Las células B y las células T son las clases principales de linfocitos y derivan de células madre hematopoyéticas pluripotenciales de la médula ósea.[57] Las células B están involucradas en la respuesta inmunitario humoral, mientras que las células T lo están en la respuesta inmunitaria mediada por células.
Las células B y T contienen moléculas receptoras que reconocen objetivos o blancos específicos. Las células T reconocen un objetivo no-propio, como un patógeno, solo después de que los antígenos (pequeños fragmentos del patógeno) han sido procesados y presentados en combinación con un receptor propio, una molécula del llamado complejo mayor de histocompatibilidad (CMH). Hay dos subtipos principales de células T: la célula T asesina (linfocito T-CD8) y la célula T colaboradora o ayudante (linfocito T-CD4). Las células T asesinas solo reconocen antígenos acoplados a moléculas del CMH de clase I, mientras que las células T colaboradoras solo reconocen antígenos acoplados a moléculas del CMH de clase II. Estos dos mecanismos de presentación de antígenos reflejan los diferentes cometidos de los dos tipos de células T. Un tercer subtipo menor lo forman las células T γ δ (células T gamma/delta), que reconocen antígenos intactos que no están acoplados a receptores CMH.[72]
Por el contrario, el receptor específico de antígeno de las células B es una molécula de anticuerpo en la superficie de la célula B, y reconoce patógenos completos sin la necesidad de que los antígenos sean procesados previamente. Cada linaje de células B expresa en su superficie un anticuerpo diferente, de forma que el conjunto completo de receptores de antígenos de las células B de un organismo, representa todos los anticuerpos que ese organismo es capaz de fabricar.[57]
Linfocitos T citotóxicos
Los linfocitos T citotóxicos, son un subgrupo de células T que matan células infectadas con virus (y otros patógenos), o que estén dañadas o enfermas por otras causas.[73] Al igual que las células B, cada tipo de célula T reconoce un antígeno diferente. Las células T asesinas son activadas cuando su receptor de células T (TCR) se liga a su antígeno específico en un complejo con el receptor del CMH de clase I de otra célula. El reconocimiento de este complejo CMH-antígeno se ve favorecido por un correceptor en la célula T, llamado CD8 (de ahí deriva su nombre T-CD8). Así, la célula T viaja a través del organismo en busca de células donde los receptores del CMH de clase I lleven este antígeno.
Cuando una célula T activada toma contacto con tales células, libera citotoxinas que forman poros en la membrana plasmática de la célula diana o receptora, permitiendo que iones, agua y toxinas entren en ella. Esto provoca el estallido de la célula diana o que experimente apoptosis.[74] La muerte de células huésped inducida por las células T asesinas tiene una gran importancia para evitar la replicación de los virus. La activación de las células T tiene unos controles muy estrictos y por lo general requiere una señal muy fuerte de activación por parte del complejo CMH/antígeno, o señales de activación adicionales proporcionadas por las células T colaboradoras (ver más abajo).[74]
Linfocitos T colaboradores
Los linfocitos T colaboradores regulan tanto la respuesta inmunitaria innata como la adaptativa, y contribuyen a determinar qué tipo de respuesta inmunitaria ofrecerá el cuerpo ante un patógeno particular.[75][76] Estos linfocitos no tienen ningún tipo de actividad citotóxica y no matan las células infectadas ni eliminan patógenos directamente. En cambio, controlan la respuesta inmunitaria dirigiendo otras células para que lleven a cabo estas tareas.
Los linfocitos T colaboradores expresan receptores de los linfocitos T que reconocen antígenos unidos a moléculas de MHC de clase II. El complejo MHC-antígeno también es reconocido por el correceptor CD4 del linfocito T colaborador, que recluta moléculas dentro del linfocito T (como la Lkc) que son responsables de la activación de dicho linfocito. Los linfocitos T colaboradores tienen una asociación más débil con el complejo MHC-antígeno que la de los linfocitos T citotóxicos, lo que significa que muchos receptores (unos 200 a 300) del linfocito T colaborador deben quedar unidos a un MHC-antígeno para activar el linfocito, mientras que los linfocitos T citotóxicos pueden ser activados por el acoplamiento de una única molécula de MHC-antígeno. La activación de los colaboradores también requiere una unión de duración superior con una célula presentadora de antígeno.[77] La activación de un linfocito T colaborador en reposo hace que libere citoquinas que influyen en la actividad de muchos tipos de células. Las señales de citocinas producidas por los linfocitos T colaboradores mejoran la función microbicida de los macrófagos y la actividad de los linfocitos T citotóxicos.[26] Además, la activación de los linfocitos T colaboradores provoca un aumento de las moléculas que se expresan en la superficie del linfocito T, como el ligando CD40 (también llamado CD154), que envía señales estimulantes adicionales requeridas generalmente para activar los linfocitos B, productores de anticuerpos.[78]
Células T γ δ
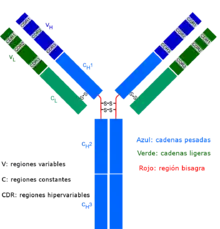
Las células T γ δ representan una pequeña subpoblación de células T caracterizada por poseer en su superficie un receptor de célula T (RCT) diferente. La mayoría de las células T tienen un RCT compuesto de dos cadenas de glucoproteínas denominadas cadenas α y β; sin embargo, en las células T γδ su receptor está formado por dos cadenas denominadas γ y δ. Este grupo de células T es, en general, menos numeroso que el de las αβ y es en la mucosa del intestino donde se las encuentra en mayor número, formando parte de una población de linfocitos denominada "linfocitos intraepiteliales".
Se desconoce en gran medida cuáles son las moléculas antigénicas que estimulan a las células T γδ, sin embargo, estas células son peculiares en el sentido de que parece que no necesitan que los antígenos sean procesados y presentados unidos a moléculas del CMH, aunque algunas reconocen a moléculas del CMH de clase IB. Por otra parte, se cree que las células T γδ desempeñan un papel principal en el reconocimiento de antígenos de naturaleza lipídica.
Las células T γδ comparten las características de las células T colaboradoras, las citotóxicas y las asesinas naturales. Al igual que otras subpoblaciones de células T no convencionales que portan RCTs invariables o constantes, como algunos subtipos de células T asesinas naturales, las γδ se encuentran en la frontera entre la inmunidad innata y la adaptativa.[80][81] Por una parte las células γδ forman parte de la inmunidad adaptativa porque son capaces de reorganizar los genes de sus RCTs para producir una diversidad de receptores y desarrollar una memoria fenotípica, es decir, ser portadoras de receptores adaptados a antígenos o patógenos concretos. Por otra parte también forman parte del sistema inmunitario innato, ya que las diferentes subpoblaciones también poseen receptores capaces de actuar como receptores de reconocimiento de patrones. Así, por ejemplo, un gran número de células T Vγ9/Vδ2 humanas (un subtipo de células comunes no peptídicas producidas por microorganismos, mientras que otro subtipo de células T, las Vδ1 en los epitelios, responden ante células epiteliales que porten indicadores de que han sufrido algún tipo de estrés.[82]
Anticuerpos y linfocitos B
El linfocito B identifica los patógenos cuando los anticuerpos de su superficie se unen a antígenos foráneos específicos.[83] Este complejo antígeno/anticuerpo pasa al interior del linfocito B donde es procesado por proteólisis y descompuesto en péptidos. El linfocito B muestra entonces estos antígenos peptídicos en su superficie unidos a moléculas del CMH de clase II. Esta combinación de CMH/antígeno atrae a un linfocito T colaborador que tenga receptores complementarios de ese complejo CMH/antígeno. La célula T libera entonces linfoquinas (el tipo de citoquinas producido por los linfocitos) y activa así al linfocito B.[84]
Cuando el linfocito B ha sido activado comienza a dividirse y su descendencia segrega millones de copias del anticuerpo que reconoce a ese antígeno. Estos anticuerpos circulan en el plasma sanguíneo y en la linfa, se ligan a los patógenos que portan esos antígenos, dejándolos marcados para su destrucción por la activación del complemento o al ser ingeridos por los fagocitos. Los anticuerpos también pueden neutralizar ciertas amenazas directamente, ligándose a toxinas bacterianas o interfiriendo con los receptores que virus y bacterias emplean para infectar las células.[85]
Sistema inmunitario adaptativo alternativo
Aunque las moléculas clásicas del sistema inmunitario adaptativo (por ejemplo, anticuerpos y receptores de células T) existen solamente en los vertebrados mandibulados, se ha descubierto una molécula diferente, y derivada de linfocitos, en vertebrados primitivos sin mandíbula, como la lamprea y animales marinos de la familia Myxinidae. Estos animales poseen una gran variedad de moléculas llamadas receptores linfocíticos variables (RLVs) que, como los receptores de antígenos de los vertebrados con mandíbula, son producidos por un número pequeño de genes (uno o dos). Se cree que estas moléculas se ligan a antígenos de los patógenos de un modo similar a como lo hacen los anticuerpos y con el mismo grado de especificidad.[86]
Memoria inmunitaria
Cuando las células B y T son activadas y comienzan a replicarse, algunos de sus descendientes se convertirán en células de memoria con un largo periodo de vida.[87] A lo largo de la vida de un Homo sapiens, estas células recordarán cada patógeno específico que se hayan encontrado y pueden desencadenar una fuerte respuesta si detectan de nuevo a ese patógeno concreto.[87] Esto es «adaptativo» porque ocurre durante el tiempo de vida de un individuo como una adaptación a una infección por ese patógeno y prepara al sistema inmunitario para futuros desafíos. La memoria inmunitaria puede ser pasiva y de corta duración o activa y de larga duración.[87]
Inmunidad pasiva

La inmunidad pasiva es generalmente de corta duración, desde unos pocos días a algunos meses. Los recién nacidos no han tenido una exposición previa a los microbios y son particularmente vulnerables a las infecciones. La madre les proporciona varias capas de protección pasiva. Durante el embarazo, un tipo particular de anticuerpo, llamado IgG, es transportado de la madre al bebé directamente a través de la placenta, así los bebés humanos tienen altos niveles de anticuerpos ya desde el nacimiento y con el mismo rango de especificidad contra antígenos que su madre.[88] La leche materna también contiene anticuerpos que al llegar al intestino del bebé le protegen de infecciones hasta que este pueda sintetizar sus propios anticuerpos.[89]
Todo esto es una forma de inmunidad pasiva porque el feto, en realidad, no fabrica células de memoria ni anticuerpos, solo los toma prestados de la madre. En medicina, la inmunidad protectora pasiva puede ser también transferida artificialmente de un individuo a otro a través de suero rico en anticuerpos.[90]
Inmunidad activa e inmunización
La memoria activa de larga duración es adquirida después de la infección, por la activación de las células T y B. La inmunidad activa puede ser también generada artificialmente, a través de la vacunación. El principio en que se basa la vacunación (también llamada inmunización) consiste en introducir un antígeno de un patógeno para estimular al sistema inmunitario y desarrollar inmunidad específica contra ese patógeno particular sin causar la enfermedad asociada con ese microorganismo.[26]
Casi todas las vacunas virales están basadas en virus «vivos» atenuados, mientras que las vacunas bacterianas están basadas en componentes o fragmentos no celulares de bacterias, incluyendo componentes inofensivos de toxinas.[26] Dado que muchas vacunas derivadas de antígenos acelulares no inducen una respuesta adaptativa lo suficientemente fuerte, a la mayoría de vacunas bacterianas se les añaden coadyuvantes que activan las células del sistema inmunitario innato presentadoras de antígenos para potenciar la inmunogenicidad.[91]
Esta inducción deliberada de una respuesta inmunitaria es efectiva porque explota la especificidad natural del sistema inmunitario, así como su inducibilidad. Siendo la enfermedad infecciosa una de las causas más frecuentes de muerte en la población humana, la vacunación representa la manipulación más eficaz del sistema inmunitario que ha desarrollado la humanidad.[57][92] Aunque cabe aclarar que no por esto las personas no se van a enfermar.
Además de brindar protección a quienes se aplican la vacuna, el acto de inmunización en la población genera un fenómeno que se conoce como «inmunidad de rebaño» o «inmunidad de grupo». Dicho efecto se logra a partir de un 95 % de las personas vacunadas. La inmunidad de grupo ayuda a proteger a los grupos más vulnerables de la sociedad como los bebés, adultos mayores, pacientes inmunocomprometidos y trasplantados. Por lo tanto, las personas vacunadas actúan como una barrera, y no permiten que los microorganismos patógenos lleguen a los más indefensos, ayudando a erradicar enfermedades, en su mayoría mortales.[93]
Trastornos de la inmunidad humana
El sistema inmunitario es un complejo notablemente eficaz que incorpora especificidad, inducibilidad y adaptación. No obstante, a veces se producen fallos que pueden agruparse, de forma genérica, dentro de las tres siguientes categorías: inmunodeficiencia, autoinmunidad e hipersensibilidad.
Inmunodeficiencias
La inmunodeficiencia ocurre cuando uno o más de los componentes del sistema inmunitario quedan inactivos. La capacidad del sistema inmunitario de responder a patógenos y enfermedades es reducida tanto en los niños como en los ancianos, y la respuesta inmunitaria empieza a entrar en declive a partir de aproximadamente los cincuenta años de edad, debido a la inmunosenescencia[94][95] (es una disminución progresiva de la respuesta inmune que afecta a todos los componentes del sistema inmunológico). En los países desarrollados, la obesidad, el alcoholismo y el uso de drogas son causas habituales de una función inmunitaria pobre.[95] Sin embargo, la malnutrición es la causa más habitual de inmunodeficiencia en los países en desarrollo.[95] Se asocia una dieta carente de suficientes proteínas con deficiencias en la inmunidad celular, la actividad del complemento, el funcionamiento de los fagocitos, las concentraciones de anticuerpos IgA y la producción de citocinas. La deficiencia de nutrientes concretos como hierro, cobre, zinc, selenio, vitaminas A, C, E y B6, y ácido fólico (vitamina B9) también reducen la respuesta inmunitaria.[95] Además, la pérdida del timo a una edad temprana a causa de una mutación genética o la extirpación quirúrgica resulta en una grave inmunodeficiencia y una gran vulnerabilidad a las infecciones.[96]
La inmunodeficiencia puede ser heredada o adquirida.[26] La enfermedad granulomatosa crónica, en que los fagocitos tienen una capacidad reducida de destruir patógenos, es un ejemplo de inmunodeficiencia heredada o congénita. El sida y algunos tipos de cáncer causan una inmunodeficiencia adquirida.[97][98]
Autoinmunidad
Las respuestas inmunes exageradas abarcan el otro extremo de la disfunción inmunitaria, particularmente las enfermedades autoinmunes. Aquí el sistema inmunitario falla en distinguir adecuadamente lo propio de lo extraño y ataca a partes del propio organismo. En circunstancias normales, muchas células T y anticuerpos reaccionan con péptidos del propio organismo.[99] Existen, sin embargo, células especializadas (localizadas en el timo y en la médula ósea) que participan en la eliminación de linfocitos jóvenes que reaccionan contra antígenos propios, para prevenir así la autoinmunidad.[83] Las reacciones autoinmunes pueden desencadenarse de varias maneras:
- Una sustancia corporal que, por lo regular, abarca un área específica y es liberada en la circulación general; y en consecuencia, se encuentra escondida en el sistema inmunitario.
- La alteración de una sustancia corporal.
- El sistema inmunitario responde a una sustancia extraña —antígeno— que parece tener las mismas características a una sustancia natural del cuerpo e involuntariamente procede a atacar tanto las sustancias del cuerpo como las extrañas.
- El mal funcionamiento de las células que controlan la producción de anticuerpos.
Hipersensibilidad
La hipersensibilidad es una inmunorespuesta que daña los tejidos propios del cuerpo. Está dividida en cuatro clases (Tipos I-IV) basándose en los mecanismos involucrados y el tiempo de desarrollo de la reacción hipersensible. El tipo I de hipersensibilidad es una reacción inmediata o anafiláctica, relacionada con alergias. Los síntomas van desde un malestar suave hasta la muerte. El tipo I de hipersensibilidad está mediado por la inmunoglobulina E, que es liberada por mastocitos y basófilos.[100] El tipo II de hipersensibilidad se produce cuando los anticuerpos se ligan a antígenos localizados sobre las células propias del paciente, marcándolas para su destrucción. También recibe el nombre de hipersensibilidad dependiente de anticuerpos o citotóxica y es mediada por anticuerpos de tipo IgG e IgM.[100] Los inmunocomplejos (agregados de antígenos, proteínas del complemento, y anticuerpos IgG e IgM) depositados en varios tejidos desencadenan la hipersensibilidad de tipo III.[100] La hipersensibilidad de tipo IV (también conocida como "hipersensibilidad de tipo retardado") generalmente tarda entre dos y tres días en desarrollarse. Las reacciones de tipo IV están implicadas en muchas enfermedades autoinmunes e infecciosas, pero también incluyen dermatitis de contacto. Estas reacciones son mediadas por las células T, monocitos y macrófagos.[100]
Otros mecanismos de defensa del huésped
Es probable que el sistema inmunitario adaptativo y de múltiples componentes surgiera con los primeros vertebrados, ya que en los invertebrados no se producen linfocitos ni respuestas humorales basadas en anticuerpos.[13] Muchas especies, sin embargo, utilizan mecanismos que parecen ser los precursores de estas funciones de la inmunidad de los vertebrados. Los sistemas inmunitarios aparecen incluso en las formas de vida más simples, como las bacterias, que utilizan un único mecanismo de defensa llamado "sistema de restricción y modificación" para protegerse de patógenos víricos llamados bacteriófagos.[101]
Los receptores de reconocimiento de patrón son proteínas que emplean casi todos los organismos para identificar moléculas relacionadas con patógenos microbianos. Los péptidos antimicrobianos llamados defensinas constituyen un componente de la respuesta inmunitario innata que se ha conservado a lo largo de la evolución, está presente en todos los animales y plantas y representa la forma principal de inmunidad sistémica de los invertebrados.[13] El sistema del complemento y las células fagocitarias también se encuentran presentes en la mayoría de los invertebrados. Las ribonucleasas y la ruta de interferencia de ARN se conservan en todos los eucariotas y se piensa que desempeñan una función en la respuesta inmunitario ante los virus y otros materiales genéticos extraños.[102]
A diferencia de los animales, las plantas no poseen células con capacidad fagocítica y la respuesta inmunitaria de la mayoría de las plantas comprende mensajeros químicos sistémicos que se distribuyen por toda la planta.[103] Cuando una parte de un vegetal resulta infectada, la planta genera una respuesta de hipersensibilidad localizada mediante la que las células del lugar de la infección sufren una rápida apoptosis para prevenir que la infección se extienda a otras partes de la planta. La resistencia sistémica adquirida (SAR) es un tipo de respuesta de las plantas que convierte a toda la planta en resistente a un agente infeccioso en particular.[103] Los mecanismos de silenciamiento de ARN tienen una especial importancia en esta respuesta sistémica, ya que pueden bloquear la replicación de virus.[104]
Inmunología de tumores

Otra función importante del sistema inmunitario es la de identificar y eliminar células tumorales. Las células transformadas de los tumores expresan antígenos que no aparecen en células normales. El sistema inmunitario considera a estos antígenos como extraños, lo que ocasiona que las células inmunitarias ataquen a las células tumorales transformadas. Los antígenos expresados por los tumores pueden tener varios orígenes;[106] algunos derivan de virus oncógenos como el papilomavirus humano, que ocasiona cáncer de cuello uterino,[107] mientras que otros son proteínas propias del organismo que se presentan en bajos niveles en células normales, pero que alcanzan altos niveles en células tumorales. Un ejemplo es una enzima llamada tirosinasa que, cuando se expresa en altos niveles, transforma a ciertas células de la piel (melanocitos) en tumores llamados melanomas.[108][109]
La principal respuesta del sistema inmunitario es destruir las células anormales por medio de células T asesinas, algunas veces con asistencia de células T colaboradoras.[109][110] Los antígenos tumorales son presentados unidos a moléculas del CMH de clase I, de forma similar a lo que ocurre con los antígenos víricos. Esto permite a las células T asesinas reconocer a las células tumorales como anormales.[111] Las células T asesinas naturales también matan células tumorales de una forma similar, especialmente si la célula tumoral tiene sobre su superficie menos moléculas del CMH de clase I de lo normal; algo que resulta habitual en los tumores.[112] A veces se generan anticuerpos contra las células tumorales, lo que permite que sean destruidas por el sistema del complemento.[106][113][114]
No obstante, algunas células tumorales evaden la acción del sistema inmunitario y generan cánceres.[115] Un mecanismo empleado a veces por las células tumorales, para evadir su detección por parte de las células T asesinas, consiste en reducir el número de moléculas del CMH de clase I en su superficie.[111] Algunas células tumorales también liberan productos que inhiben la respuesta inmunitaria, por ejemplo al secretar la citoquina TGF-β, la cual suprime la actividad de macrófagos y linfocitos.[116] Además, también puede desarrollarse tolerancia inmunológica frente a los antígenos tumorales, de forma que el sistema inmunitario deja de atacar a las células tumorales.[115]
Regulación fisiológica
Las hormonas pueden modular la sensibilidad del sistema inmunitario. Por ejemplo, se sabe que las hormonas sexuales femeninas estimulan las reacciones tanto del sistema inmunitario adaptativo[117] como del innato.[118] Algunas enfermedades autoinmunes como el lupus eritematoso afectan con mayor frecuencia a las mujeres, y su comienzo coincide a menudo con la pubertad. Por el contrario, andrógenos como la testosterona parece que deprimen al sistema inmunitario.[119] Otras hormonas, como la prolactina y la hormona de crecimiento o vitaminas como la vitamina D, parece que también regulan las respuestas del sistema inmunitario.[120][121] Se piensa que el descenso progresivo en los niveles de hormonas con la edad, pudiera ser parcialmente responsable del debilitamiento de las respuestas inmunitarias en individuos de edad avanzada.[122] A la inversa, algunas hormonas son reguladas por el sistema inmunitario, sobre todo la actividad de la hormona tiroidea.[123]
El sistema inmunitario se ve potenciado con el sueño y el descanso,[124] mientras que resulta perjudicado por el estrés.[125] Las dietas pueden afectar al sistema inmunitario; por ejemplo frutas frescas, vegetales y comida rica en ciertos ácidos grasos favorecen el mantenimiento de un sistema inmunitario saludable.[126] Asimismo, la desnutrición fetal puede causar una debilitación de por vida del sistema inmunitario.[127] En las medicinas tradicionales, se cree que algunas plantas pueden estimular el sistema inmunitario y ciertos estudios así lo han sugerido,[128] aunque su mecanismo de acción es complejo y difícil de caracterizar.
Sueño y descanso
El sistema inmunológico se ve afectado por el sueño y el descanso, y la falta de sueño es perjudicial para la función inmunitaria.[129] Los circuitos de retroalimentación complejos que involucran citocinas, como la IL-1 y el TNF-α producidos en respuesta a la infección, también parecen desempeñar un papel en la regulación del sueño con movimientos oculares no rápidos (REM).[130] Por lo tanto, la respuesta inmunitaria a la infección puede provocar cambios en el ciclo del sueño, incluido un aumento del sueño de ondas lentas en relación con el sueño REM.[131]
En las personas que sufren de privación del sueño, las inmunizaciones activas pueden tener un efecto disminuido y pueden resultar en una menor producción de anticuerpos y una menor respuesta inmune, de lo que se observaría en una persona bien descansada.[132] Además, proteínas como NFIL3, que se ha demostrado que están estrechamente entrelazadas tanto con la diferenciación de células T como con los ritmos circadianos, pueden verse afectadas por la alteración de los ciclos naturales de luz y oscuridad a través de casos de privación del sueño. Estas alteraciones pueden provocar un aumento de afecciones crónicas como enfermedades cardíacas, dolor crónico y asma.[133]
Además de las consecuencias negativas de la privación del sueño, se ha demostrado que el sueño y el sistema circadiano entrelazado tienen fuertes efectos reguladores sobre las funciones inmunológicas que afectan tanto a la inmunidad innata como a la adaptativa. Primero, durante la etapa temprana del sueño de ondas lentas, una caída repentina de los niveles sanguíneos de cortisol, epinefrina y norepinefrina provoca un aumento de los niveles sanguíneos de las hormonas leptina, hormona del crecimiento hipofisario y prolactina. Estas señales inducen un estado proinflamatorio a través de la producción de citocinas proinflamatorias IL-1, IL-12, TNF-alfa e interferón gamma. Estas citocinas luego estimulan funciones inmunes como la activación, proliferación y diferenciación de las células inmunes. Durante este tiempo de una respuesta inmune adaptativa que evoluciona lentamente, hay un pico en las células indiferenciadas o menos diferenciadas, como las células T de memoria central y vírgenes. Además de estos efectos, el medio de las hormonas producidas en este momento (leptina, hormona del crecimiento hipofisario y prolactina) respalda las interacciones entre las células presentadoras de antígenos (APC) y las células T, un cambio del equilibrio de citocinas Th1 / Th2 hacia uno que respalda a Th1, un aumento de la proliferación general de células Th y migración de células T vírgenes a los ganglios linfáticos. También se cree que esto favorece la formación de una memoria inmunitaria duradera mediante el inicio de respuestas inmunitarias Th1.[134]
Durante los períodos de vigilia, las células efectoras diferenciadas, como las células NK y los linfocitos T citotóxicos, alcanzan su punto máximo para provocar una respuesta eficaz contra cualquier patógeno intruso. Las moléculas antiinflamatorias, como el cortisol y las catecolaminas, también alcanzan su punto máximo durante los momentos activos de vigilia. La inflamación causaría graves deterioros cognitivos y físicos si ocurriera durante las horas de vigilia, y la inflamación puede ocurrir durante las horas de sueño debido a la presencia de melatonina. La inflamación causa una gran cantidad de estrés oxidativo y la presencia de melatonina durante las horas de sueño podría contrarrestar activamente la producción de radicales libres durante este tiempo.
Manipulación en la medicina

La respuesta inmunitaria puede ser manipulada para suprimir respuestas no deseadas de la autoinmunidad, la alergia y el rechazo de trasplantes, así como para estimular respuestas protectoras contra patógenos que en gran medida eluden la acción del sistema inmunitario. Se emplean fármacos inmunosupresores para controlar las enfermedades autoinmunes o la inflamación cuando produce grandes daños en los tejidos, o para prevenir el rechazo de un órgano trasplantado.[57][135]
Los fármacos antiinflamatorios se emplean para controlar los efectos de la inflamación. Los corticosteroides son los más poderosos de estos medicamentos; sin embargo, tienen muchos efectos tóxicos colaterales y su uso debe ser controlado estrictamente.[136] Por ello, a menudo, se emplean dosis más bajas de antiinflamatorios junto con fármacos inmunosupresores y citotóxicos como el metotrexato o la azatioprina. Los fármacos citotóxicos inhiben la inmunorespuesta destruyendo células que se están dividiendo, como las células T que han sido activadas. Sin embargo, la destrucción es indiscriminada, por lo que otros órganos y tipos de células resultan afectados, lo que ocasiona efectos colaterales.[135] Los fármacos inmunodepresores como la ciclosporina evitan que las células T respondan correctamente a las señales, inhibiendo rutas de transducción de señales.[137]
Los fármacos de mayor peso molecular (> 500 dalton) pueden provocar la neutralización de la respuesta inmunitaria, particularmente si son suministrados repetidamente, o en dosis grandes. Esto limita la eficacia de los fármacos constituidos por grandes péptidos y proteínas (que generalmente superan los 6000 dalton). En algunos casos, el fármaco no es inmunógeno en sí mismo, pero puede ser coadministrado con un medicamento inmunógeno, como el Taxol. Se han desarrollado métodos computacionales para predecir la inmunogenicidad de péptidos y proteínas, que resultan particularmente útiles en el diseño de anticuerpos terapéuticos, la valoración de la probable virulencia de las mutaciones que afecten a partículas víricas de recubrimiento y la validación de nuevos fármacos basados en péptidos. Las primeras técnicas se basaban principalmente en el hecho observado de que los aminoácidos hidrófilos se encuentran presentes, en mayor cantidad que los aminoácidos hidrófobos, en los epítopos (determinantes antigénicos que producen una interacción específica reversible con una inmunoglobulina y consisten en un grupo de aminoácidos localizados sobre la superficie del antígeno);[138] sin embargo, más recientemente se han empleado técnicas de Aprendizaje Automático, que se sirven de bases de datos de epítopos conocidos, generalmente de proteínas víricas bien estudiadas.[139] Se ha creado una base de datos de acceso público para la catalogación de epítopos de patógenos que se sabe son reconocidos por células B.[140] Los estudios de inmunogenicidad basados en la bioinformática, constituyen un campo emergente que se conoce con el nombre de inmunoinformática.[141]
Manipulación por los patógenos
El éxito de cualquier patógeno depende de su habilidad para eludir las respuestas inmunitarias del huésped. Por ello, los patógenos han desarrollado diferentes métodos que les permiten infectar con éxito al huésped, al mismo tiempo que evaden la destrucción producida por la inmunidad.[142] Las bacterias frecuentemente logran sobrepasar las barreras físicas al secretar enzimas que digieren la barrera —por ejemplo, utilizando un sistema de secreción de tipo II—.[143] Alternativamente, al usar un sistema de secreción tipo III, pueden insertar un tubo hueco en la célula huésped que les provee de un conducto para trasladar proteínas del patógeno al huésped; las proteínas transportadas por el tubo son utilizadas frecuentemente para desarmar las defensas del huésped.[144]
Una estrategia utilizada por varios patógenos para eludir al sistema inmunitario innato es la replicación intracelular (también llamada patogénesis intracelular). En ella, un patógeno pasa la mayor parte de su ciclo vital dentro de células huésped en donde se protege del contacto directo con células inmunitarias, anticuerpos y proteínas del complemento. Algunos ejemplos de patógenos intracelulares incluyen virus, bacterias del género Salmonella causantes de toxiinfecciones alimentarias y los parásitos eucariotas que causan la malaria (Plasmodium falciparum) y la leismaniosis (Leishmania spp.). Otras bacterias, como el Mycobacterium tuberculosis, viven dentro de una cápsula protectora que evita su lisis por el complemento.[145] Muchos patógenos secretan componentes que disminuyen o desvían la respuesta inmunitaria del huésped.[142] Algunas bacterias forman biopelículas para protegerse de las células y proteínas del sistema inmunitario. Estas biopelículas están presentes en muchas infecciones que cursan con éxito, como por ejemplo las infecciones crónicas producidas por Pseudomonas aeruginosa y Burkholderia cenocepacia características de la Fibrosis quística.[146] Otras bacterias generan proteínas de superficie que se ligan a los anticuerpos, volviéndolos ineficaces. Como ejemplos se pueden citar: estreptococos (proteína G), Staphylococcus aureus (proteína A), y Peptostreptococcus magnus (proteína L).[147]
Los mecanismos empleados por los virus para eludir al sistema inmunitario adaptativo son más complejos. El enfoque más sencillo consiste en cambiar rápidamente los epítopos no esenciales (aminoácidos o azúcares) de la superficie del invasor, mientras se mantienen los epítopos esenciales ocultos. El VIH, por ejemplo, muta regularmente las proteínas de su envoltura viral que le son esenciales para entrar en las células huésped que son su objetivo. Estos cambios frecuentes en antígenos pueden explicar el hecho de no haber logrado producir vacunas dirigidas contra estas proteínas.[148] Otra estrategia común para evitar ser detectados por el sistema inmunitario consiste en enmascarar sus antígenos con proteínas de la célula huésped. Así, en el VIH, la envoltura que recubre al virión está formada por la membrana más externa de la célula huésped; tales virus «autocamuflados» dificultan que el sistema inmunitario los identifique como algo no propio.[149]
Historia de la inmunología
La inmunología es una ciencia que examina la estructura y función del sistema inmunitario. Se origina en la medicina y en los primeros estudios sobre las causas de la inmunidad a las enfermedades. La referencia más antigua a la inmunidad se produce durante la plaga de Atenas en el 430 a. C., donde Tucídides notó que algunas personas que se habían recuperado de un brote anterior de la enfermedad podían atender a los enfermos sin contraer la enfermedad por segunda vez.[150] Esta observación de inmunidad adquirida fue luego utilizada por Louis Pasteur en el desarrollo de la vacunación y en su teoría microbiana de la enfermedad.[151] La teoría de Pasteur se oponía a las teorías contemporáneas sobre las enfermedades, tales como la teoría miasmática. No se confirmó que los microorganismos fueran la causa de las enfermedades infecciosas hasta 1891, cuando Robert Koch enunció sus postulados, por los que recibió el Premio Nobel en 1905.[152] En 1901, con el descubrimiento del virus de la fiebre amarilla por Walter Reed, se confirmó que los virus son patógenos humanos.[153]
Se produjo un gran avance en la inmunología hacia el final del siglo XIX, gracias al rápido desarrollo de los estudios de inmunidad humoral y de inmunidad celular.[154] De particular importancia fue el trabajo de Paul Ehrlich, quien propuso la Teoría de la cadena lateral para explicar la especificidad de la reacción antígeno-anticuerpo; sus contribuciones al entendimiento de la inmunología humoral fueron reconocidos con el Premio Nobel en 1908, recibido en conjunto con Elie Metchnikoff, el fundador de la inmunología celular.[155]
Peter Gorer descubrió en 1936 el antígeno H-2 del ratón, y consigo el primer complejo mayor de histocompatibilidad (MHC). Mientras tanto, Peter Medawar y Thomas Gibson pudieran aclarar funciones importantes de las células inmunitarias. En 1948, Astrid Fagraeus descubrió que los anticuerpos son producidos por los linfocitos B del plasma. Un año más tarde, Frank Macfarlane Burnet y Frank Fenner publicaron su hipótesis sobre la tolerancia inmunitaria, que sería confirmada algunos años más tarde por Jacques Miller con el descubrimiento de la eliminación de linfocitos T autorreactivos en el timo. En 1957, Frank Macfarlane Burnet describió la teoría de la selección clonal como principio central de la inmunidad adaptiva.[156]
A finales de la década de 1960 y principios de la década de 1970, John David y Barry Bloom descubrieron el Factor Inhibidor de Migración de los Macrófagos (MIF) y una nueva clase de sustancias secretadas por los linfocitos. Dudley Dumonde acuñó el término "linfocina" para estas sustancias. Stanley Cohen, que en 1986 consiguió el Premio Nobel de Fisiología o Medicina por su descubrimiento de los factores de crecimiento NGF y EGF,[157][158] comenzó a estudiar a principios de la década de 1970 las funciones de los factores denominados "linfocinas" junto con Takeshi Yoshida. Descubrieron que estas sustancias pertenecen a un grupo de sustancias mensajeras que son producidas por muchos tipos diferentes de células del sistema inmunitario. En 1974 Stanley Cohen propuso el término "citocina", que se consolidó con el descubrimiento de más sustancias de este tipo. Desde entonces se han descubierto más de cien nuevas citocinas, la estructura y las funciones de las cuales han sido investigadas en detalle.
Véase también
Referencias
- F. Lozano Soto (2012). «Introducción al sistema inmunológico, sus principales elementos y la respuesta inmunitaria». En Farreras; Rozman, eds. Medicina Interna (17 edición). Barcelona, España: Elsevier España. pp. 2453-2488. ISBN 9788480868969.
- Delvis, Peter J. (2016). «Panorama general del sistema inmunitario» (en inglés). Merck Sharp & Dohme Corp. Consultado el 21 de junio de 2016.
- Berkow, Roberts (2008). «16». Manual Merck: Home edition (2da. edición). Océano. p. 837. ISBN 84-494-1184-X.
- National Institute of Allergy and Infectious Diseases, ed. (diciembre de 2013). «Overview of the Immune System» [Descripción general del sistema inmunitario] (en inglés). Consultado el 21 de junio de 2016.
- National Institute of Allergy and Infectious Diseases, ed. (2003). Understanding the Immune System: How It Works [Entendiendo el sistema inmune: cómo trabaja] (en inglés). Estados Unidos. Consultado el 21 de junio de 2016.
- Middleton D, Curran M, Maxwell L (August de 2002). «Natural killer cells and their receptors». Transplant Immunology 10 (2-3): 147-64. PMID 12216946. doi:10.1016/S0966-3274(02)00062-X.
- Pancer Z, Cooper MD (2006). «The evolution of adaptive immunity». Annual Review of Immunology 24: 497-518. PMID 16551257. doi:10.1146/annurev.immunol.24.021605.090542.
- Berkow, Roberts (2008). «16». Manual Merck: Home edition (2da. edición). Océano. p. 838. ISBN 84-494-1184-X.
- Rus H, Cudrici C, Niculescu F (2005). «The role of the complement system in innate immunity». Immunologic Research 33 (2): 103-12. PMID 16234578. doi:10.1385/IR:33:2:103.
- Matzinger P (April de 2002). «The danger model: a renewed sense of self». Science 296 (5566): 301-5. PMID 11951032. doi:10.1126/science.1071059.
- Berkow, Roberts (2008). «16». Manual Merck: Home edition (2da. edición). Océano. p. 838. ISBN 84-494-1184-X.
- Agerberth B, Gudmundsson GH (2006). «Host antimicrobial defence peptides in human disease». Current Topics in Microbiology and Immunology 306: 67-90. PMID 16909918. doi:10.1007/3-540-29916-5_3.
- Beck, Gregory; Gail S. Habicht (noviembre de 1996). «Immunity and the Invertebrates» (PDF). Scientific American: 60-66. Consultado el 18 de octubre de 2009.
- Berkow, Roberts (2008). «16». Manual Merck: Home edition (2da. edición). Océano. p. 842. ISBN 84-494-1184-X.
- Berkow, Roberts (2008). «16». Manual Merck: Home edition (2da. edición). Océano. p. 846. ISBN 84-494-1184-X.
- Joos L, Tamm M (2005). «Breakdown of pulmonary host defense in the immunocompromised host: cancer chemotherapy». Proceedings of the American Thoracic Society 2 (5): 445-8. PMID 16322598. doi:10.1513/pats.200508-097JS.
- Copeland KF, Heeney JL (December de 1996). «T helper cell activation and human retroviral pathogenesis». Microbiological Reviews 60 (4): 722-42. PMC 239461. PMID 8987361.
- «Diccionario médico-biológico, histórico y etimológico: inmunidad». Ediciones Universidad Salamanca. febrero de 2012. Archivado desde el original el 1 de agosto de 2016. Consultado el 21 de junio de 2016.
- «Sistema inmunitario y no sistema inmunológico». Fundéu BBVA. 3 de octubre de 2011. Consultado el 21 de junio de 2016.
- Gomila, Xavier Albons; Puigventós, Àngels Egea. «Immunològic i immunitari: un esforç il·lògic?» [Inmunológico e inmunitario: ¿un esfuerzo ilógico?]. Assessorament Lingüístic i Terminologia (en catalán) (Barcelona, España: Serveis Lingüístics, Universitat de Barcelona). Consultado el 21 de junio de 2016.
- http://www.ugr.es/~eianez/inmuno/cap_02.htm
- Litman G, Cannon J, Dishaw L (2005). «Reconstructing immune phylogeny: new perspectives.». Nat Rev Immunol 5 (11): 866-79. PMID 16261174.
- Mayer, Gene (2006). «Immunology - Chapter One: Innate (non-specific) Immunity». Microbiology and Immunology On-Line Textbook. USC School of Medicine. Archivado desde el original el 16 de febrero de 2007. Consultado el 1 de enero de 2007.
- Smith A.D. (Ed) Oxford dictionary of biochemistry and molecular biology. (1997) Oxford University Press. ISBN 0-19-854768-4
- Alberts, Bruce; Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, and Peter Walters (2002). Molecular Biology of the Cell; Fourth Edition. New York and London: Garland Science. ISBN 0-8153-3218-1.
- Reid G, Jass J, Sebulsky M, McCormick J (2003). «Potential uses of probiotics in clinical practice». Clin Microbiol Rev 16 (4): 658-72. PMID 14557292. doi:10.1128/CMR.16.4.658-672.2003.
- Medzhitov R (2007). «Recognition of microorganisms and activation of the immune response». Nature 449 (7164): 819-26. PMID 17943118. doi:10.1038/nature06246.
- Boyton R, Openshaw P (2002). «Pulmonary defences to acute respiratory infection». Br Med Bull 61: 1-12. PMID 11997295. doi:10.1093/bmb/61.1.1.
- Agerberth B, Gudmundsson G. «Host antimicrobial defence peptides in human disease.». Curr Top Microbiol Immunol 306: 67-90. PMID 16909918.
- Moreau J, Girgis D, Hume E, Dajcs J, Austin M, O'Callaghan R (2001). «Phospholipase A(2) in rabbit tears: a host defense against Staphylococcus aureus.». Invest Ophthalmol Vis Sci 42 (10): 2347-54. PMID 11527949.
- Hankiewicz J, Swierczek E (1974). «Lysozyme in human body fluids.». Clin Chim Acta 57 (3): 205-9. PMID 4434640.
- Fair W, Couch J, Wehner N (1976). «Prostatic antibacterial factor. Identity and significance.». Urology 7 (2): 169-77. PMID 54972.
- Yenugu S, Hamil K, Birse C, Ruben S, French F, Hall S (2003). «Antibacterial properties of the sperm-binding proteins and peptides of human epididymis 2 (HE2) family; salt sensitivity, structural dependence and their interaction with outer and cytoplasmic membranes of Escherichia coli.». Biochem J 372 (Pt 2): 473-83. PMID 12628001.
- Gorbach S (1990). «Lactic acid bacteria and human health». Ann Med 22 (1): 37 - 41. PMID 2109988.
- Hill L, Embil J (1986). «Vaginitis: current microbiologic and clinical concepts.». CMAJ 134 (4): 321-31. PMID 3510698.
- Salminen S, Gueimonde M, Isolauri E (2005). «Probiotics that modify disease risk». J Nutr 135 (5): 1294 - 8. PMID 15867327.
- Reid G, Bruce A (2003). «Urogenital infections in women: can probiotics help?». Postgrad Med J 79 (934): 428-32. PMID 12954951. doi:10.1136/pmj.79.934.428.
- Karakitsos D, Karabinis A (September de 2008). «Hypothermia therapy after traumatic brain injury in children». N. Engl. J. Med. 359 (11): 1179-80. PMID 18788094.
- Axelrod YK, Diringer MN (May de 2008). «Temperature management in acute neurologic disorders». Neurol Clin 26 (2): 585-603, xi. PMID 18514828. doi:10.1016/j.ncl.2008.02.005.
- Capítulo 58 en: Walter F., PhD. Boron (2003). Medical Physiology: A Cellular And Molecular Approaoch. Elsevier/Saunders. p. 1300. ISBN 1-4160-2328-3.
-
- Rhoades, R. and Pflanzer, R. Human physiology, third edition, chapter 27 Regulation of body temperature, p. 820 Clinical focus: pathogenesis of fever. ISBN 0-03-005159-2
- Laupland KB (July de 2009). «Fever in the critically ill medical patient». Crit. Care Med. 37 (7 Suppl): S273-8. PMID 19535958. doi:10.1097/CCM.0b013e3181aa6117.
- Fauci, Anthony, et al. (2008). Harrison's Principles of Internal Medicine (17 edición). McGraw-Hill Professional. pp. 117-121. ISBN 9780071466332.
- Schaffner A. Fever—useful or noxious symptom that should be treated? Ther Umsch 2006; 63: 185-8. PMID 16613288
- Soszynski D. The pathogenesis and the adaptive value of fever. Postepy Hig Med Dosw 2003; 57: 531-54. PMID 14737969
- Craven, R and Hirnle, C. (2006). Fundamentals of nursing: Human health and function. Fourth edition. p. 1044
- Lewis, SM, Heitkemper, MM, and Dirksen, SR. (2007). Medical-surgical nursing: Assessment and management of clinical problems. sixth edition. p. 212
- «Fever». Medline Plus Medical Encyclopedia. U.S. National Library of Medicine. Consultado el 20 de mayo de 2009.
- Kawai T, Akira S (2006). «Innate immune recognition of viral infection». Nat Immunol 7 (2): 131-7. PMID 16424890.
- Campbell, Neil A.; Reece, Jane B. (2007). Biología. Ed. Médica Panamericana. ISBN 978-84-7903-998-1. Consultado el 30 de abril de 2020.
- Miller, SB (2006). «Prostaglandins in Health and Disease: An Overview». Seminars in Arthritis and Rheumatism 36 (1): 37-49. PMID 16887467.
- Ogawa Y, Calhoun WJ. (2006). «The role of leukotrienes in airway inflammation.». J Allergy Clin Immunol. 118 (4): 789-98. PMID 17030228.
- Le Y, Zhou Y, Iribarren P, Wang J (2004). «Chemokines and chemokine receptors: their manifold roles in homeostasis and disease». Cell Mol Immunol 1 (2): 95-104. PMID 16212895.
- Martin P, Leibovich S (2005). «Inflammatory cells during wound repair: the good, the bad and the ugly.». Trends Cell Biol 15 (11): 599-607. PMID 16202600.
- Mayer, Gene (2006). «Immunology - Chapter Two: Complement». Microbiology and Immunology On-Line Textbook. USC School of Medicine. Archivado desde el original el 9 de febrero de 2007. Consultado el 1 de enero de 2007.
- Janeway CA, Jr. et al (2005). Immunobiology. (6th ed. edición). Garland Science. ISBN 0-443-07310-4.
- Liszewski M, Farries T, Lublin D, Rooney I, Atkinson J. «Control of the complement system.». Adv Immunol 61: 201-83. PMID 8834497.
- Sim R, Tsiftsoglou S (2004). «Proteases of the complement system.». Biochem Soc Trans 32 (Pt 1): 21-7. PMID 14748705.
- Ryter A (1985). «Antimicrobial functions of mononuclear phagocytes». Comp Immunol Microbiol Infect Dis 8 (2): 119-33. PMID 3910340.
- Langermans J, Hazenbos W, van Furth R (1994). «Antimicrobial functions of mononuclear phagocytes». J Immunol Methods 174 (1-2): 185-94. PMID 8083520.
- May R, Machesky L (2001). «Phagocytosis and the actin cytoskeleton». J Cell Sci 114 (Pt 6): 1061-77. PMID 11228151.
- Salzet M, Tasiemski A, Cooper E (2006). «Innate immunity in lophotrochozoans: the annelids». Curr Pharm Des 12 (24): 3043-50. PMID 16918433.
- Zen K, Parkos C (2003). «Leukocyte-epithelial interactions». Curr Opin Cell Biol 15 (5): 557-64. PMID 14519390.
- Stvrtinová, Viera; Ján Jakubovský and Ivan Hulín (1995). Inflammation and Fever from Pathophysiology: Principles of Disease. Computing Centre, Slovak Academy of Sciences: Academic Electronic Press. Archivado desde el original el 11 de julio de 2001. Consultado el 1 de enero de 2007.
- Bowers, William (2006). «Immunology -Chapter Thirteen: Immunoregulation». Microbiology and Immunology On-Line Textbook. USC School of Medicine. Archivado desde el original el 16 de diciembre de 2006. Consultado el 4 de enero de 2007.
- Guermonprez P, Valladeau J, Zitvogel L, Théry C, Amigorena S. «Antigen presentation and T cell stimulation by dendritic cells». Annu Rev Immunol 20: 621-67. PMID 11861614.
- Krishnaswamy G, Ajitawi O, Chi D. «The human mast cell: an overview.». Methods Mol Biol 315: 13-34. PMID 16110146.
- Kariyawasam H, Robinson D (2006). «The eosinophil: the cell and its weapons, the cytokines, its locations». Semin Respir Crit Care Med 27 (2): 117-27. PMID 16612762.
- Middleton D, Curran M, Maxwell L (2002). «Natural killer cells and their receptors». Transpl Immunol 10 (2-3): 147-64. PMID 12216946.
- Pancer Z, Cooper M. «The evolution of adaptive immunity». Annu Rev Immunol 24: 497-518. PMID 16551257.
- Holtmeier W, Kabelitz D. «gammadelta T cells link innate and adaptive immune responses». Chem Immunol Allergy 86: 151-83. PMID 15976493.
- Harty J, Tvinnereim A, White D. «CD8+ T cell effector mechanisms in resistance to infection». Annu Rev Immunol 18: 275-308. PMID 10837060.
- Radoja S, Frey A, Vukmanovic S (2006). «T-cell receptor signaling events triggering granule exocytosis». Crit Rev Immunol 26 (3): 265-90. PMID 16928189.
- Abbas A, Murphy K, Sher A (1996). «Functional diversity of helper T lymphocytes». Nature 383 (6603): 787-93. PMID 8893001. doi:10.1038/383787a0.
- McHeyzer-Williams L, Malherbe L, McHeyzer-Williams M (2006). «Helper T cell-regulated B cell immunity». Curr Top Microbiol Immunol 311: 59-83. PMID 17048705. doi:10.1007/3-540-32636-7_3.
- Kovacs B, Maus M, Riley J, Derimanov G, Koretzky G, June C, Finkel T (2002). «Human CD8+ T cells do not require the polarization of lipid rafts for activation and proliferation». Proc Natl Acad Sci U S a 99 (23): 15006-11. PMID 12419850. doi:10.1073/pnas.232058599.
- Grewal I, Flavell R (1998). «CD40 and CD154 in cell-mediated immunity». Annu Rev Immunol 16: 111-35. PMID 9597126. doi:10.1146/annurev.immunol.16.1.111.
- «Understanding the Immune System: How it Works» (PDF) (en inglés). National Institute of Allergy and Infectious Diseases. Consultado el 15 de febrero de 2013.
- Girardi M (2006). «Immunosurveillance and immunoregulation by γδ T cells». J Invest Dermatol 126 (1): 25-31. PMID 16417214.
- Holtmeier W, Kabelitz D (2005). «gammadelta T cells link innate and adaptive immune responses». Chemical Immunology and Allergy 86: 151-83. PMID 15976493. doi:10.1159/000086659.
- Holtmeier W, Kabelitz D (2005). «γδ T cells link innate and adaptive immune responses». Chem Immunol Allergy 86: 151-183. PMID 15976493.
- Sproul T, Cheng P, Dykstra M, Pierce S (2000). «A role for MHC class II antigen processing in B cell development». Int Rev Immunol 19 (2-3): 139-55. PMID 10763706.
- Kehry M, Hodgkin P (1994). «B-cell activation by helper T-cell membranes». Crit Rev Immunol 14 (3-4): 221-38. PMID 7538767.
- Bowers, William (2006). «Immunology - Chapter nine: Cells involved in immune responses». Microbiology and Immunology On-Line Textbook. USC School of Medicine. Archivado desde el original el 16 de diciembre de 2006. Consultado el 4 de enero de 2007.
- M.N. Alder, I.B. Rogozin, L.M. Iyer, G.V. Glazko, M.D. Cooper, Z. Pancer (2005). «Diversity and Function of Adaptive Immune Receptors in a Jawless Vertebrate». Science 310 (5756): 1970 - 1973. PMID 16373579.
- Berkow, Roberts (2008). «16». Manual Merck: Home edition (2da. edición). Océano. p. 842. ISBN 84-494-1184-X.
- Saji F, Samejima Y, Kamiura S, Koyama M (1999). «Dynamics of immunoglobulins at the feto-maternal interface.». Rev Reprod 4 (2): 81-9. PMID 10357095. Archivado desde el original el 24 de junio de 2008. Consultado el 1 de marzo de 2007.
- Van de Perre P (2003). «Transfer of antibody via mother's milk.». Vaccine 21 (24): 3374-6. PMID 12850343.
- Keller, Margaret A. and E. Richard Stiehm (2000). «Passive Immunity in Prevention and Treatment of Infectious Diseases.». Clinical Microbiology Reviews 13 (4): 602-614. PMID 11023960.
- Singh M, O'Hagan D (1999). «Advances in vaccine adjuvants». Nat Biotechnol 17 (11): 1075-81. PMID 10545912.
- Death and DALY estimates for 2002 by cause for WHO Member States. Organización Mundial de la Salud. Visitado el 1 de enero de 2007.
- Esteban, Pablo (1508291938). «“Vacunarse es un acto solidario” | Guadalupe Nogués, una bióloga dedicada a la comunicación y la educación en ciencias». PAGINA12. Consultado el 30 de abril de 2020.
- Aw D, Silva A, Palmer D (2007). «Immunosenescence: emerging challenges for an ageing population». Immunology 120 (4): 435-446. PMID 17313487. doi:10.1111/j.1365-2567.2007.02555.x.
- Chandra, RK (1997). «Nutrition and the immune system: an introduction». American Journal of Clinical Nutrition. Vol 66: 460S-463S. PMID 9250133. Free full-text pdf available
- Miller JF (2002). «The discovery of thymus function and of thymus-derived lymphocytes». Immunol. Rev. 185: 7-14. PMID 12190917. Archivado desde el original el 15 de diciembre de 2018. Consultado el 27 de mayo de 2010.
- Joos L, Tamm M (2005). «Breakdown of pulmonary host defense in the immunocompromised host: cancer chemotherapy». Proc Am Thorac Soc 2 (5): 445-8. PMID 16322598. doi:10.1513/pats.200508-097JS.
- Copeland K, Heeney J (1996). «T helper cell activation and human retroviral pathogenesis». Microbiol Rev 60 (4): 722-42. PMID 8987361.
- Miller J (1993). «Self-nonself discrimination and tolerance in T and B lymphocytes». Immunol Res 12 (2): 115-30. PMID 8254222.
- Ghaffar, Abdul (2006). «Immunology - Chapter Seventeen: Hypersensitivity Reactions». Microbiology and Immunology On-Line Textbook. USC School of Medicine. Archivado desde el original el 16 de diciembre de 2006. Consultado el 1 de enero de 2007.
- Bickle T, Krüger D (1993). «Biology of DNA restriction». Microbiol Rev 57 (2): 434-50. PMID 8336674.
- Stram Y, Kuzntzova L. (2006). «Inhibition of viruses by RNA interference». Virus Genes 32 (3): 299-306. PMID 16732482.
- Schneider, David (Spring 2005). «Innate Immunity - Lecture 4: Plant immune responses». Stanford University Department of Microbiology and Immunology. Archivado desde el original el 9 de junio de 2007. Consultado el 1 de enero de 2007.
- Baulcombe D (2004). «RNA silencing in plants». Nature 431 (7006): 356-63. PMID 15372043.
- Morgan R et al. (2006). «Cancer regression in patients after transfer of genetically engineered lymphocytes». Science 314: 126-129. PMID 16946036.
- Andersen MH, Schrama D, Thor Straten P, Becker JC (2006). «Cytotoxic T cells». J Invest Dermatol 126 (1): 32-41. PMID 16417215.
- Boon T, van der Bruggen P (1996). «Human tumor antigens recognized by T lymphocytes». J Exp Med 183: 725-29. PMID 8642276.
- Castelli C, Rivoltini L, Andreola G, Carrabba M, Renkvist N, Parmiani G (2000). «T cell recognition of melanoma-associated antigens». J Cell Physiol 182: 323-31. PMID 10653598.
- Romero P, Cerottini JC, Speiser DE (2006). «The human T cell response to melanoma antigens». Adv Immunol. 92: 187-224. PMID 17145305.
- Gerloni M, Zanetti M. (2005). «CD4 T cells in tumor immunity». Springer Semin Immunopathol 27 (1): 37-48. PMID 15965712.
- Seliger B, Ritz U, Ferrone S (2006). «Molecular mechanisms of HLA class I antigen abnormalities following viral infection and transformation». Int J Cancer 118 (1): 129-38. PMID 16003759.
- Hayakawa Y, Smyth MJ. (2006). «Innate immune recognition and suppression of tumors». Adv Cancer Res 95: 293-322. PMID 16860661.
- Guevara-Patino JA, Turk MJ, Wolchok JD, Houghton AN (2003). «Immunity to cancer through immune recognition of altered self: studies with melanoma». Adv Cancer Res. 90: 157-77. PMID 14710950.
- Renkvist N, Castelli C, Robbins PF, Parmiani G (2001). «A listing of human tumor antigens recognized by T cells». Cancer Immunol Immunother 50: 3-15. PMID 11315507.
- Seliger B (2005). «Strategies of tumor immune evasion». BioDrugs 19 (6): 347-54. PMID 16392887.
- Frumento G, Piazza T, Di Carlo E, Ferrini S (2006). «Targeting tumor-related immunosuppression for cancer immunotherapy». Endocr Metab Immune Disord Drug Targets 6 (3): 233-7. PMID 17017974.
- Wira, CR; Crane-Godreau M, Grant K (2004). «Endocrine regulation of the mucosal immune system in the female reproductive tract». En In: Ogra PL, Mestecky J, Lamm ME, Strober W, McGhee JR, Bienenstock J (eds.), ed. Mucosal Immunology. San Francisco: Elsevier. ISBN 0-12-491543-4.
- Lang, TJ (2004). «Estrogen as an immunomodulator». Clin Immunol 113: 224-230. PMID 15507385.
Moriyama, A; Shimoya K, Ogata I et al. (1999). «Secretory leukocyte protease inhibitor (SLPI) concentrations in cervical mucus of women with normal menstrual cycle». Molecular Human Reproduction 5: 656-661. PMID 10381821.
Cutolo, M; Sulli A, Capellino S, Villaggio B, Montagna P, Seriolo B, Straub RH (2004). «Sex hormones influence on the immune system: basic and clinical aspects in autoimmunity». Lupus 13: 635-638. PMID 15485092.
King, AE; Critchley HOD, Kelly RW (2000). «Presence of secretory leukocyte protease inhibitor in human endometrium and first trimester decidua suggests an antibacterial role». Molecular Human Reproduction 6: 191-196. PMID 10655462. - Fimmel, S; Zouboulis CC (2005). «Influence of physiological androgen levels on wound healing and immune status in men». Aging Male 8: 166-174. PMID 16390741.
- Dorshkind, K; Horseman ND (2000). «The Roles of Prolactin, Growth Hormone, Insulin-Like Growth Factor-I, and Thyroid Hormones in Lymphocyte Development and Function: Insights from Genetic Models of Hormones and Hormone Receptor Deficiency». Endocrine Reviews 21: 292-312. PMID 10857555. Archivado desde el original el 31 de julio de 2010. Consultado el 1 de marzo de 2007.
- Nagpal, Sunil; Songqing Naand and Radhakrishnan Rathnachalam (2005). «Noncalcemic Actions of Vitamin D Receptor Ligands». Endocrine Reviews 26 (5): 662-687. PMID 15798098. Archivado desde el original el 19 de febrero de 2009. Consultado el 1 de marzo de 2007.
- Hertoghe, T (2005). «The “multiple hormone deficiency” theory of aging: Is human senescence caused mainly by multiple hormone deficiencies?». Annals of the New York Academy of Science 1051: 448-465. PMID 16399912.
- Klein, JR (2006). «The immune system as a regulator of thyroid hormone activity». Exp Biol Med 231: 229-236. PMID 16514168.
- Lange, T; Perras B, Fehm HL, Born J (2003). «Sleep Enhances the Human Antibody response to Hepatitis A Vaccination». Psychosomatic Medicine 65: 831-835. PMID 14508028.
- Khansari, DN; Murgo AJ, Faith RE (1990). «Effects of stress on the immune system». Immunology Today 11: 170-175. PMID 2186751.
- Pond, CM (2005). «Adipose tissue and the immune system». Prostaglandins, Leukotrienes, and Essential Fatty Acids 73: 17-30. PMID 15946832.
- Langley-Evans, SC; Carrington LJ (2006). «Diet and the developing immune system». Lupus 15: 746-752. PMID 17153845.
- Spelman, K; Burns J, Nichols D, Winters N, Ottersberg S, Tenborg M (2006). «Modulation of cytokine expression by traditional medicines: a review of herbal immunomodulators». Alternative Medicine reviews 11: 128-150. PMID 16813462.
Brush, J; Mendenhall E, Guggenheim A, Chan T, Connelly E, Soumyanth A, Buresh R, Barrett R, Zwickey H (2006). «The effect of Echinacea purpurea, Astragalus membranaceus and Glycyrrhiza glabra on CD69 expression and immune cell activation in humans». Phytotherapy Research 20: 687-695. PMID 16807880. - Bryant PA, Trinder J, Curtis N (Jun 2004). «Sick and tired: Does sleep have a vital role in the immune system?». Nature Reviews. Immunology 4 (6): 457-67. PMID 15173834. S2CID 29318345. doi:10.1038/nri1369.
- Krueger JM, Majde JA (May 2003). «Humoral links between sleep and the immune system: research issues». Annals of the New York Academy of Sciences 992 (1): 9-20. Bibcode:2003NYASA.992....9K. PMID 12794042. S2CID 24508121. doi:10.1111/j.1749-6632.2003.tb03133.x.
- Majde JA, Krueger JM (Dec 2005). «Links between the innate immune system and sleep». The Journal of Allergy and Clinical Immunology 116 (6): 1188-98. PMID 16337444. doi:10.1016/j.jaci.2005.08.005.
- Taylor DJ, Kelly K, Kohut ML, Song KS (2017). «Is Insomnia a Risk Factor for Decreased Influenza Vaccine Response?». Behavioral Sleep Medicine 15 (4): 270-287. PMC 5554442. PMID 27077395. doi:10.1080/15402002.2015.1126596.
- Krueger JM (2008). «The role of cytokines in sleep regulation». Current Pharmaceutical Design 14 (32): 3408-16. PMC 2692603. PMID 19075717. doi:10.2174/138161208786549281.
- Besedovsky L, Lange T, Born J (Jan 2012). «Sleep and immune function». Pflügers Archiv 463 (1): 121-37. PMC 3256323. PMID 22071480. doi:10.1007/s00424-011-1044-0.
- Taylor A, Watson C, Bradley J (2005). «Immunosuppressive agents in solid organ transplantation: Mechanisms of action and therapeutic efficacy». Crit Rev Oncol Hematol 56 (1): 23-46. PMID 16039869.
- Barnes P (2006). «Corticosteroids: the drugs to beat». Eur J Pharmacol 533 (1-3): 2-14. PMID 16436275.
- Masri M (2003). «The mosaic of immunosuppressive drugs». Mol Immunol 39 (17-18): 1073-7. PMID 12835079.
- Welling GW, Wiejer WJ, van der Zee R, Welling-Werster S. (1985). «Prediction of sequential antigenic regions in proteins». J Mol Recognit 88 (2): 215-8. PMID 2411595.
- Sollner J, Mayer B. (2006). Machine learning approaches for prediction of linear B-cell epitopes on proteins. 19 (3). pp. 200-8. PMID 16598694.
- Saha S, Bhasin M, Raghava GP. (2005). «Bcipep: a database of B-cell epitopes.». BMC Bioinformatics 6 (1): 79. PMID 15921533.
- Flower DR, Doytchinova IA. (2002). «Immunoinformatics and the prediction of immunogenicity.». Appl Bioinformatics 1 (4): 167-76. PMID 15130835.
- Finlay B, McFadden G (2006). «Anti-immunology: evasion of the host immune system by bacterial and viral pathogens». Cell 124 (4): 767-82. PMID 16497587.
- Cianciotto NP. (2005). «Type II secretion: a protein secretion system for all seasons». Trends Microbiol. 13 (12): 581-8. PMID 16216510.
- Winstanley C, Hart CA (2001). «Type III secretion systems and pathogenicity islands». J Med Microbiol. 50 (2): 116-26. PMID 11211218.
- Finlay B, Falkow S (1997). «Common themes in microbial pathogenicity revisited». Microbiol Mol Biol Rev 61 (2): 136-69. PMID 9184008.
- Kobayashi H (2005). «Airway biofilms: implications for pathogenesis and therapy of respiratory tract infections». Treat Respir Med 4 (4): 241-53. PMID 16086598.
- Housden N, Harrison S, Roberts S, Beckingham J, Graille M, Stura E, Gore M (2003). «Immunoglobulin-binding domains: Protein L from Peptostreptococcus magnus». Biochem Soc Trans 31 (Pt 3): 716-8. PMID 12773190.
- Burton, Dennis R.; Robyn L. Stanfield and Ian A. Wilson (2005). «Antibody vs. HIV in a clash of evolutionary titans». Proc Natl Acad Sci U S A. 102 (42): 14943-8. PMID 16219699.
- Cantin R, Methot S, Tremblay MJ. (2005). «Plunder and stowaways: incorporation of cellular proteins by enveloped viruses». J Virol. 79 (11): 6577-87. PMID 15890896.
- Retief F, Cilliers L (1998). «The epidemic of Athens, 430-426 BC». S Afr Med J 88 (1): 50-3. PMID 9539938.
- Plotkin S (2005). «Vaccines: past, present and future». Nat Med 11 (4 Suppl): S5-11. PMID 15812490.
- El Premio Nobel de Medicina de 1905 Nobelprize.org Visitado 8 de enero de 2007 (en inglés).
- Mayor Walter Reed, Cuerpo médico del ejército de Estados Unidos Walter Reed Army Medical Center. Visitado el 8 de enero de 2007.
-
Metchnikoff, Elie; Translated by F.G. Binnie. (1905). Immunity in Infective Diseases (Versión in extenso: Google Books). Cambridge University Press. ISBN 68025143
|isbn=incorrecto (ayuda). - El Premio Nobel de Medicina de 1908 Nobelprize.org Visitado el 8 de enero de 2007
- Forsdyke, D. R. (1995). "The Origins of the Clonal Selection Theory of Immunity" Archivado el 30 de julio de 2012 en Wayback Machine. FASEB. Journal 9:164-66
- Shampo, M A; Kyle R A (Juny de 1999). «Stanley Cohen--Nobel laureate for growth factor». Mayo Clin. Proc. (Estados Unidos) 74 (6): 600. ISSN 0025-6196. PMID 10377936.
- El Premio Nobel de Fisiología o Medicina 1986 Nobelprize.org Consultado el 28 de julio de 2011.
Enlaces externos
 Wikimedia Commons alberga una categoría multimedia sobre Sistema inmunitario.
Wikimedia Commons alberga una categoría multimedia sobre Sistema inmunitario.- Anatomía y fisiología - sistema inmunitario.
- Enciclopedia Médica en español - Respuesta inmunitaria.
 Datos: Q1059
Datos: Q1059- Multimedia: Immune system / Q1059