Betacoronavirus
Betacoronavirus (ou β-coronavirus) est l'un des quatre genres connus de coronavirus. Il est classé dans la sous-famille des Orthocoronavirinae, la famille des Coronaviridae et l'ordre des Nidovirales. Il regroupe des virus à ARN simple brin enveloppés, de sens positif, d'origine zoonotique.
.jpg.webp)
Sous-genres de rang inférieur
Le genre Betacoronavirus est organisé en 5 sous-genres et une quinzaine d'espèces. Dans la littérature plus ancienne, ce genre est également connu sous le nom de « coronavirus du groupe 2 ».
Les espèces de Betacoronavirus de la plus haute importance clinique concernant les humains sont Betacoronavirus 1 (OC43) et Human coronavirus HKU1 dans le sous-genre Embecovirus, SARS-CoV et SARS-CoV-2 dans le sous-genre Sarbecovirus[2] et MERS-CoV dans le sous-genre Merbecovirus[3].
Virologie
Les bétacoronavirus (β-CoV) infectent principalement les chauves-souris[4], créant des lignées dérivées virales distinctes réputées affecter les voies respiratoires et/ou entériques d'autres espèces de mammifères[4], dont l'humain (avec HCoV-HKU1 et HCoV-OC43). Chez les animaux, des β-CoV différents ont à ce jour été trouvés chez des espèces aussi différentes que le chameau ou dromadaire et le lapin[5],[6],[7],[8], et aussi chez le porc (PHEV), le cheval (coronavirus équin, ECoV), et le chien (coronavirus respiratoire canin, CRCoV).
Par ailleurs, plusieurs virus liés au genre Betacoronavirus (virus « de type β-CoV », tous actuellement inclus dans l'espèce unique Betacoronavirus-1), ont été retrouvés dans les voies digestives (entériques) et/ou respiratoires de ruminants domestiques et sauvages : chez des ruminants domestiqués tels qu'ovins et caprins[9],[10], le buffle d'eau (Bubalus bubalis)[11], le lama (Lama glama) et l'alpaga (Vicugna pacos)[12],[13]. Six espèces de cervidés en portent (caribou/renne (Rangifer tarandus caribou), le cerf élaphe/wapiti (Cervus elaphus), le sambar (Cervus unicolor), le cerf de Virginie (Odocoileus virginianus), le cerf sika (Cervus nippon yesoensis) et le cerf d'eau (Hydropotes inermis)[14]. La girafe (Giraffa camelopardalis)[15], plusieurs antilopes[16],[17], le bison d'Europe (Bison bonasus), le thar d'Himalaya (Hemitragus jemlahicus)[17], et le dromadaire (Camelus dromedarius)[18].
Chez l'humain, les bétacoronavirus humains sont des agents du rhume commun. Certains bétacoronavirus ont provoqué des épidémies humaines, avec généralement de la fièvre et des symptômes respiratoires. Ils incluent :
- deux souches de SARSr-CoV :
- le SARS-CoV, agent du syndrome respiratoire aigu sévère (SRAS, 2003) ;
- le SARS-CoV-2 (deux lignées « L » et « S ») agent de la pandémie de COVID-19 (2019-2021) ;
- MERS-CoV, agent du syndrome respiratoire du Moyen-Orient (MERS, 2012).
Gestion écoépidémiologique du risque
Risque pandémique : depuis le début des années 2000, de nombreux écologues, virologues et vétérinaires ont alerté sur le fait que les conditions d'une pandémie zoonotique grave étaient réunies. Au vu de la liste d'espèces présentée ci-dessus, et sachant que les bétacoronavirus (β-CoV) sont des archétypes de virus franchissant facilement les barrières interspécifiques[4], les chasseurs, éleveurs et profession liées aux abattoirs et marchés humides en première ligne en termes de contact avec un éventuel virus émergeant. Selon ces deux infectiologues, une surveillance génomique « massive » est pour cela nécessaire dans la faune sauvages (et pas que pour les CoV), de même qu'un séquençage massif et partagé des souches de SARS-CoV-2 détectées dans la faune et chez les patients ayant développé une COVID-19, ceci afin de comprendre l'origine de la COVID-19, et éviter d'autres pandémies similaires ou plus graves. Avant cela il est urgent ;
- de fermer tous les marchés humides, et mettre en place une gestion plus respectueuse de l'environnement[4] ;
- comprendre les interactions entre les CoV et leurs hôtes, in vitro (cultures cellulaires, explants ex-vivo des voies respiratoires) et in vivo (étude d'animaux sensibles à l'infection par le SARS-CoV-2)[4] ;
- préparer de nouveaux médicaments anti-coronaviraux sans attendre une prochaine épidémie ou pandémie[4].
L'OMS et l'OIE, sous l'égide de l'ONU recommandent de traiter les pandémies zoonotiques via une approche globale et balistique dite « One Health »[19].
Classification
Le genre Betacoronavirus de la sous-famille des Orthocoronavirinae de la famille des Coronaviridae comprend 5 sous-genres reconnus par l'International Committee on Taxonomy of Viruses (ICTV)[20],[21] :
| β-CoV[22] |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Virions
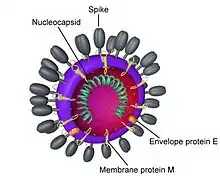
Les particules virales, quasi-sphériques, sont généralement couvertes de grandes projections de surface (~ 20 nm) en forme de pétale ou de pointe (les "spikes" en anglais), qui créent une image qui rappelle la couronne solaire en micrographie électronique: cette propriété est à l'origine du nom des coronavirus. Ces « pointes », des protéines S (spiculaires) présentes à la surface du virus, déterminent le tropisme de l'hôte.
Le sous-genre Embecovirus diffère des autres en ce que les virions ont une protéine en forme de pointe plus courte appelée hémagglutinine estérase (HE).
Plusieurs structures des protéines spiculaires ont été résolues. Le domaine de liaison au récepteur dans la protéine S (péplomère) des alpha- et bétacoronavirus est catalogué comme InterPro: IPRO018548[32],[33]. Les protéines S s'assemblent en trimères (PDB: 3jcl[34],[35], 6acg[36],[37]) dont la structure centrale rappelle celle des protéines F (fusion) de paramyxovirus[38].
Notes et références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Betacoronavirus » (voir la liste des auteurs).
- ICTV. International Committee on Taxonomy of Viruses. Taxonomy history. Published on the Internet https://talk.ictvonline.org/., consulté le 24 janvier 2021
- « Phylogeny of SARS-like betacoronaviruses », nextstrain (consulté le )
- Memish, Zumla, Al-Hakeem et Al-Rabeeah, « Family Cluster of Middle East Respiratory Syndrome Coronavirus Infections », New England Journal of Medicine, vol. 368, no 26, , p. 2487–94 (PMID 23718156, DOI 10.1056/NEJMoa1303729)
- (en) Nicola Decaro et Alessio Lorusso, « Novel human coronavirus (SARS-CoV-2): A lesson from animal coronaviruses », sur Veterinary Microbiology, (PMID 32402329, PMCID PMC7195271, DOI 10.1016/j.vetmic.2020.108693, consulté le ), p. 108693
- Woo, Wang, Lau et Xu, « Comparative analysis of twelve genomes of three novel group 2c and group 2d coronaviruses reveals unique group and subgroup features », Journal of Virology, vol. 81, no 4, , p. 1574–85 (PMID 17121802, PMCID 1797546, DOI 10.1128/JVI.02182-06)
- Lau, Woo, Yip et Fan, « Isolation and characterization of a novel Betacoronavirus subgroup A coronavirus, rabbit coronavirus HKU14, from domestic rabbits », Journal of Virology, vol. 86, no 10, , p. 5481–96 (PMID 22398294, PMCID 3347282, DOI 10.1128/JVI.06927-11)
- Lau, Poon, Wong et Wang, « Coexistence of different genotypes in the same bat and serological characterization of Rousettus bat coronavirus HKU9 belonging to a novel Betacoronavirus subgroup », Journal of Virology, vol. 84, no 21, , p. 11385–94 (PMID 20702646, PMCID 2953156, DOI 10.1128/JVI.01121-10)
- Zhang, Zheng, Agwanda et Ommeh, « Serological evidence of MERS-CoV and HKU8-related CoV co-infection in Kenyan camels », Emerging Microbes & Infections, vol. 8, no 1, , p. 1528–1534 (DOI 10.1080/22221751.2019.1679610)
- Reinhardt, G., Zamora, J., Tadich, N., Polette, M., Aguilar, M., Riedemann, S., & Palisson, J. (1995). Diagnosis of coronavirus in sheep in Valdivia province, Xth Region, Chile. Archivos de Medicina Veterinaria, 27, 129-132.,
- (en) Dong-Kun Yang, In-Jin Hwang, Byoung-Han Kim et Chang-Hee Kweon, « Serosurveillance of Viral Diseases in Korean Native Goats (Capra hircus) », Journal of Veterinary Medical Science, vol. 70, no 9, , p. 977–979 (ISSN 0916-7250 et 1347-7439, DOI 10.1292/jvms.70.977, lire en ligne, consulté le )
- (en) Nicola Decaro, Vito Martella, Gabriella Elia et Marco Campolo, « Biological and genetic analysis of a bovine-like coronavirus isolated from water buffalo (Bubalus bubalis) calves », Virology, vol. 370, no 1, , p. 213–222 (PMID 17916374, PMCID PMC7103353, DOI 10.1016/j.virol.2007.08.031, lire en ligne, consulté le )
- (en) Christopher K. Cebra, Donald E. Mattson, Rocky J. Baker et Robert J. Sonn, « Potential pathogens in feces from unweaned llamas and alpacas with diarrhea », Journal of the American Veterinary Medical Association, vol. 223, no 12, , p. 1806–1808 (ISSN 0003-1488, DOI 10.2460/javma.2003.223.1806, lire en ligne, consulté le )
- (en) L. Jin, C.K. Cebra, R.J. Baker et D.E. Mattson, « Analysis of the genome sequence of an alpaca coronavirus », Virology, vol. 365, no 1, , p. 198–203 (PMID 17459444, PMCID PMC7185508, DOI 10.1016/j.virol.2007.03.035, lire en ligne, consulté le )
- (en) Haitham Mohamed Amer, « Bovine-like coronaviruses in domestic and wild ruminants », Animal Health Research Reviews, vol. 19, no 2, , p. 113–124 (ISSN 1466-2523 et 1475-2654, PMID 30683171, PMCID PMC7108644, DOI 10.1017/S1466252318000117, lire en ligne, consulté le )
- (en) Mustafa Hasoksuz, Konstantin Alekseev, Anastasia Vlasova et Xinsheng Zhang, « Biologic, Antigenic, and Full-Length Genomic Characterization of a Bovine-Like Coronavirus Isolated from a Giraffe », Journal of Virology, vol. 81, no 10, , p. 4981–4990 (ISSN 0022-538X et 1098-5514, PMID 17344285, PMCID PMC1900194, DOI 10.1128/JVI.02361-06, lire en ligne, consulté le )
- (en) Konstantin P. Alekseev, Anastasia N. Vlasova, Kwonil Jung et Mustafa Hasoksuz, « Bovine-Like Coronaviruses Isolated from Four Species of Captive Wild Ruminants Are Homologous to Bovine Coronaviruses, Based on Complete Genomic Sequences », Journal of Virology, vol. 82, no 24, , p. 12422–12431 (ISSN 0022-538X et 1098-5514, PMID 18842722, PMCID PMC2593316, DOI 10.1128/JVI.01586-08, lire en ligne, consulté le )
- (en) Joon-Yee Chung, Hye-Ryoung Kim, You-Chan Bae et O-Soo Lee, « Detection and characterization of bovine-like coronaviruses from four species of zoo ruminants », Veterinary Microbiology, vol. 148, nos 2-4, , p. 396–401 (PMID 20880639, PMCID PMC7117225, DOI 10.1016/j.vetmic.2010.08.035, lire en ligne, consulté le )
- (en) P. C. Y. Woo, S. K. P. Lau, C. S. F. Lam et A. K. L. Tsang, « Discovery of a Novel Bottlenose Dolphin Coronavirus Reveals a Distinct Species of Marine Mammal Coronavirus in Gammacoronavirus », Journal of Virology, vol. 88, no 2, , p. 1318–1331 (ISSN 0022-538X, PMID 24227844, PMCID PMC3911666, DOI 10.1128/JVI.02351-13, lire en ligne, consulté le )
- (en) Contini C et Di Nuzzo M, « The Novel Zoonotic COVID-19 Pandemic: An Expected Global Health Concern », sur Journal of infection in developing countries, (PMID 32235085, consulté le )
- (en) « International Committee on Taxonomy of Viruses (ICTV) », sur talk.ictvonline.org (consulté le )
- DOI:10.1186/s44149-021-00005-9
- DOI:10.1128/JVI.01600-13
- DOI:10.1073/pnas.0506735102
- Shin Murakami, Tomoya Kitamura, Jin Suzuki, Ryouta Sato, Toshiki Aoi, Marina Fujii, Hiromichi Matsugo, Haruhiko Kamiki, Hiroho Ishida, Akiko Takenaka-Uema, Masayuki Shimojima et Taisuke Horimoto, « Detection and Characterization of Bat Sarbecovirus Phylogenetically Related to SARS-CoV-2, Japan », Emerging Infectious Diseases, vol. 26, no 12, , p. 3025–3029 (DOI 10.3201/eid2612.203386)
- Tommy Tsan-Yuk Lam, Na Jia, Ya-Wei Zhang, Marcus Ho-Hin Shum, Jia-Fu Jiang, Hua-Chen Zhu, Yi-Gang Tong, Yong-Xia Shi, Xue-Bing Ni, Yun-Shi Liao, Wen-Juan Li, Bao-Gui Jiang, Wei Wei, Ting-Ting Yuan, Kui Zheng, Xiao-Ming Cui, Jie Li, Guang-Qian Pei, Xin Qiang, William Yiu-Man Cheung, Lian-Feng Li, Fang-Fang Sun, Si Qin, Ji-Cheng Huang, Gabriel M. Leung, Edward C. Holmes, Yan-Ling Hu, Yi Guan et Wu-Chun Cao, « Identifying SARS-CoV-2-related coronaviruses in Malayan pangolins », Nature, vol. 583, no 7815, , p. 282–285 (DOI 10.1038/s41586-020-2169-0)
- Ping Liu, Jing-Zhe Jiang, Xiu-Feng Wan, Yan Hua, Linmiao Li, Jiabin Zhou, Xiaohu Wang, Fanghui Hou, Jing Chen, Jiejian Zou et Jinping Chen, « Are pangolins the intermediate host of the 2019 novel coronavirus (SARS-CoV-2)? », PLOS Pathogens, vol. 16, no 5, , e1008421 (DOI 10.1371/journal.ppat.1008421)
- (en) Vibol Hul, Deborah Delaune, Erik A. Karlsson, Alexandre Hassanin, Putita Ou Tey, Artem Baidaliuk, Fabiana Gámbaro, Vuong Tan Tu, Lucy Keatts, Jonna Mazet, Christine Johnson, Philippe Buchy, Philippe Dussart, Tracey Goldstein, Etienne Simon-Lorière et Veasna Duong, « A novel SARS-CoV-2 related coronavirus in bats from Cambodia », sur bioRxiv, (DOI 10.1101/2021.01.26.428212), p. 2021.01.26.428212
- S Wacharapluesadee, CW Tan, P Maneeorn, P Duengkae, F Zhu, Y Joyjinda, T Kaewpom, WN Chia, W Ampoot, BL Lim, K Worachotsueptrakun, VC Chen, N Sirichan, C Ruchisrisarod, A Rodpan, K Noradechanon, T Phaichana, N Jantarat, B Thongnumchaima, C Tu, G Crameri, MM Stokes, T Hemachudha et LF Wang, « Evidence for SARS-CoV-2 related coronaviruses circulating in bats and pangolins in Southeast Asia. », Nature Communications, vol. 12, no 1, , p. 972 (PMID 33563978, PMCID 7873279, DOI 10.1038/s41467-021-21240-1)
- H Zhou, X Chen, T Hu, J Li, H Song, Y Liu, P Wang, D Liu, J Yang, EC Holmes, AC Hughes, Y Bi et W Shi, « A Novel Bat Coronavirus Closely Related to SARS-CoV-2 Contains Natural Insertions at the S1/S2 Cleavage Site of the Spike Protein. », Current biology : CB, vol. 30, no 11, , p. 2196-2203.e3 (PMID 32416074, DOI 10.1016/j.cub.2020.05.023)
- « Addendum: A pneumonia outbreak associated with a new coronavirus of probable bat origin », Nature, vol. 588, no 7836, , E6 (PMID 33199918, DOI 10.1038/s41586-020-2951-z, lire en ligne)
- (en) « Spike receptor binding domain (InterPro entry IPR018548) », sur https://www.ebi.ac.uk/interpro (consulté le )
- Huang, Qi, Lu et Wang, « Putative Receptor Binding Domain of Bat-Derived Coronavirus HKU9 Spike Protein: Evolution of Betacoronavirus Receptor Binding Motifs. », Biochemistry, vol. 55, no 43, , p. 5977-5988 (PMID 27696819, DOI 10.1021/acs.biochem.6b00790)
- (en-US) A. C. Walls, M. A. Tortorici, B. J. Bosch et B. Frenz, « Cryo-electron microscopy structure of a coronavirus spike glycoprotein trimer. », Nature, vol. 531, , p. 47–52 (DOI 10.2210/pdb3jcl/pdb, lire en ligne, consulté le )
- (en) « Cryo-electron microscopy structure of a coronavirus spike glycoprotein trimer » (consulté le )
- (en-US) W. Song, M. Gui, X. Wang et Y. Xiang, « Cryo-EM structure of the SARS coronavirus spike glycoprotein in complex with its host cell receptor ACE2. », PLoS Pathog., vol. 14, , e1007236–e1007236 (DOI 10.2210/pdb6acg/pdb, lire en ligne, consulté le )
- (en) « Trypsin-cleaved and low pH-treated SARS-CoV spike glycoprotein and ACE2 complex, ACE2-bound conformation 1 », sur Protein Data Bank (consulté le )
- Walls, Tortorici, Bosch et Frenz, « Cryo-electron microscopy structure of a coronavirus spike glycoprotein trimer », Nature, vol. 531, no 7592, , p. 114–117 (DOI 10.1038/nature16988)