Therapsida
Theromorpha • Thérapsides, Théromorphes
Ne doit pas être confondu avec Synapsida.

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Super-classe | Tetrapoda |
| Classe | Synapsida |
| Clade | Eupelycosauria |
| Clade | Sphenacodontia |
| Clade | Sphenacodontoidea |
Sous-ordres de rang inférieur
- † Raranimus
- incertae sedis :
- † Dinosaurus
- † Niaftasuchus
- † Biarmosuchia
- Eutherapsida
- † Dinocephalia
- Neotherapsida
- † Anomodontia
- Theriodontia
- † Gorgonopsia
- Eutheriodontia
- † Therocephalia
- Cynodontia
- † Divers taxons éteints
- Mammalia
Les thérapsides (Therapsida[alpha 1]), parfois appelés théromorphes (Theromorpha) selons certaines sources, forment un groupe majeur de synapsides eupélycosauriens qui comprend les mammifères et les lignées ancestrales de ces derniers. Bon nombre des traits considérés aujourd'hui comme spécifiques aux mammifères ont leurs origines chez les premiers thérapsides, y compris les membres qui sont articulés sous le corps, et non latéralement comme chez de nombreux reptiles et amphibiens.
Les plus vieux thérapsides connus sont datées de plus de 272 millions d'années et descendent des sphénacodontes, remplaçant progressivement les pélycosaures (terme non valide utilisé pour désigner les synapsides plus basaux) en tant que grands animaux terrestres dominants du Permien moyen jusqu'au Trias inférieur. À la suite de l'extinction Permien-Trias, l'importance relative des thérapsides diminue par rapport aux reptiles diapsides qui se diversifient rapidement au cours du Trias moyen, seuls les dicynodontes et les euthériodontes survivant à cet évènement, les autres lignées s'étant éteintes.
Les thérapsides euthériodontes comprennent les cynodontes, un groupe très diversifié qui donne naissance aux mammifères durant le Jurassique moyen, ces derniers étant parmi les rares représentants du groupe ayant survécu à l'extinction Trias-Jurassique. Les derniers des cynodontes non mammaliens, les tritylodontidés, continuent d'exister jusqu'au Crétacé inférieur.
Caractéristiques
Mâchoire et dentitions
Les fosses temporales des thérapsides sont plus grandes que celles des pélycosaures et leurs mâchoires sont plus complexes et puissantes : les dents se différencient en incisives frontales pour pincer, en grandes canines latérales pour perforer et déchirer ainsi que des molaires pour cisailler et hacher les aliments, en raison du fait que beaucoup d'espèces actuelles et en particulier non mammifères possèdent un palais charnu qui leur permet déjà de manger et respirer en même temps.
Posture
Les pattes des thérapsides sont positionnées plus verticalement sous leurs corps que ne le sont celles des reptiles et des pélycosaures. De plus, par rapport à ces groupes, les pieds sont plus symétriques, avec les premier et dernier orteils courts et les orteils moyens longs, une indication que l'axe du pied est placé parallèlement à celui de l'animal, sans s'étendre latéralement. Cette orientation aurait donné une démarche plus semblable à celle d'un mammifère que celle d'un lézard ou d'un pélycosaure[3].
Physiologie
La physiologie des thérapsides est mal connue. La plupart des thérapsides du Permien possèdent un foramen pinéal, ce qui indique qu'ils ont un œil pariétal comme de nombreux reptiles et amphibiens modernes. L'œil pariétal joue un rôle important dans la thermorégulation et le rythme circadien des ectothermes, mais est absent chez les mammifères modernes, qui sont endothermiques[4]. Vers la fin du Permien, les dicynodontes, les thérocéphales et les cynodontes montrent des tendances parallèles vers la perte du foramen pinéal, ce dernier étant complètement absent chez les cynodontes probainognathiens. Les preuves des isotopes de l'oxygène, qui sont corrélées à la température corporelle, suggèrent que la plupart des thérapsides du Permien sont ectothermes et que l'endothermie aurait évolué de manière convergente chez les dicynodontes et les cynodontes vers la fin du Permien[5]. En revanche, des preuves histologiques suggèrent que l'endothermie est partagée à travers les thérapsides[6], alors que les estimations du débit sanguin et de la durée de vie chez Morganucodon suggèrent que même les premiers mammaliaformes ont des taux métaboliques de type « reptilien »[7]. Des preuves de cornets respiratoires, supposées indiquer une endothermie, sont rapportées chez le thérocéphale Glanosuchus (en), mais une étude ultérieure montre que les sites de fixation apparents des cornets peuvent simplement être le résultat d'une distorsion du crâne[8].
Tégument
L'évolution des téguments chez les thérapsides est mal connue et il existe peu de fossiles qui fournissent des preuves directes de la présence ou de l'absence de fourrure. Les synapsides les plus basaux avec des preuves directes et sans ambiguïté de fourrure sont les docodontes, qui sont des mammaliaformes très étroitement liés au groupe-couronne des mammifères. La peau d'un crâne fossilisée du dinocéphale Estemmenosuchus est décrite comme montrant une peau « glandulaire », manquant à la fois d'écailles et de poils[9].
Des coprolithes contenant ce qui semble être des poils sont trouvés dans des archives fossile datant du Permien[10],[11]. Bien que la source de ces poils ne soit pas connue avec certitude, ils peuvent suggérer que les poils sont présents dans au moins certains thérapsides de cette période.
La fermeture du foramen pinéal chez les cynodontes probainognathiens peut indiquer une mutation du gène régulateur Msx2, qui est impliqué à la fois dans la fermeture du toit du crâne et dans le maintien des follicules pileux chez la souris[12]. Cela suggère que les fourrures ont peut-être d'abord évolué chez les probainognathiens, bien que cela n'exclue pas entièrement une origine plus ancienne de cette dernière[12].
Les vibrisses proviennent probablement des cynodontes probainognathiens[12],[13]. Certaines études déduisent une origine plus ancienne des vibrisses sur la base de la présence de foramens sur le museau des thérocéphales et des premiers cynodontes, mais la disposition des foramens dans ces taxons ressemble en réalité à celles de certains sauropsides[14], ce qui rend la présence de vibrisses semblables à ceux des mammifères peu probable[13].
Évolution des thérapsides
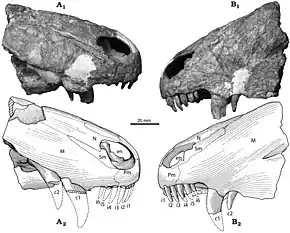
Les thérapsides descendent d'un groupe de pélycosaures appelés sphénacodontes[16],[17]. Ces derniers deviennent les animaux terrestres dominants du Permien moyen, remplaçant leurs ancêtres pélycosaures. Le taxon Therapsida se compose de quatre sous-groupes principaux : les biarmosuchiens, les dinocéphales, les anomodontes (herbivores) ainsi que les thériodontes (qui incluent notamment les ancêtres des mammifères). Après un bref sursaut de diversité évolutive, les dinocéphales s'éteignent durant la fin du Permien moyen, mais le sous-groupe d'anomodontes connu sous le nom de dicynodontes ainsi que les thériodontes tels que les gorgonopsiens et les thérocéphales prospèrent, rejoints tout à la fin du Permien par les premiers cynodontes.
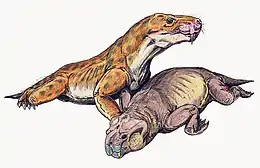
Comme tous les autres tétrapodes, les thérapsides sont gravement touchés par l'extinction Permien-Trias : les gorgonopsiens et les biarmosuchiens disparaissent complètement et les groupes restants (dicynodontes, thérocephales et cynodontes), réduits à une poignée d'espèces chacun durant le Trias inférieur. Les dicynodontes, maintenant représentés par un seul groupe de grands herbivores trapus, les Kannemeyeriiformes et les cynodontes de taille moyenne (comprenant à la fois des formes carnivores et herbivores), prospèrent dans le monde entier tout au long du Trias inférieur et moyen. Ils disparaissent des archives fossiles dans une grande partie de la Pangée à la fin du Carnien (Trias supérieur), bien qu'ils continuent d'exister quelque temps de plus dans la bande équatoriale humide et au sud.
Parmi les quelques exceptions figurent les eucynodontes, encore plus dérivés, dont aux moins trois groupes d'entre eux réussissent à survivre. Ils apparaissent à la fin du Trias et la famille extrêmement proche des mammifères, les tritylodontidés, survit jusqu'au Crétacé inférieur. Une autre famille extrêmement proche des mammifères, les trithélodontidés, est inconnue des archives fossiles datant d'après le Jurassique inférieur. Les mammaliaformes incarnent le troisième groupe, comprenant Morganucodon et des formes apparentées. Certains taxonomistes désignent ces animaux comme étant des « mammifères », bien que la plupart limitent ce terme au groupe-couronne qui contient les représentants modernes.
Certains cynodontes ne faisant pas partie des eucynodontes survivent de l'extinction Permien-Trias : Thrinaxodon, Galesaurus et Platycraniellus sont connus des archives fossiles du Trias inférieur. Cependant, au Trias moyen, seuls les eucynodontes subsistent.

Les dicynodontes font partie des groupes de thérapsides les plus avancés de la fin du Permien et prospèrent jusqu'à la fin du Trias.
Les thérocéphales, proches parents des cynodontes, réussissent également à survivre lors de cette extinction et continuent à se diversifier tout au long du Trias inférieur. Cependant, à l'approche de la fin de cette période, les thérocéphales sont en déclin jusqu'à une éventuelle extinction, probablement dépassés par la lignée des diapsides qui se diversifie rapidement, équipée de systèmes respiratoires sophistiqués mieux adaptés au monde très chaud, sec et pauvre en oxygène de la fin du Trias.
Les mammifères (incluant l'homme) sont les seuls thérapsides encore existants de nos jours. Le groupe-couronne des mammifères, dont les plus vieux représentants connus datent du Jurassique inférieur, est issu d'une radiation évolutive des mammaliaformes contenant également des formes apparentées tels que les docodontes. Les mammaliaformes eux-mêmes descendent des probainognathiens, une des deux lignées des eucynodontes.
Classification
Six grands groupes de thérapsides sont généralement reconnus : Biarmosuchia, Dinocephalia, Anomodontia, Gorgonopsia, Therocephalia et Cynodontia. Un clade unissant les thérocéphales et les cynodontes, appelé Eutheriodontia, est bien soutenu, mais les relations entre les quatre autres clades sont controversées[20]. L'hypothèse la plus largement acceptée des relations entre les thérapsides est celui d'Hopson et Barghausen, proposée pour la première fois en . Selon cette hypothèse, les biarmosuchiens sont le premier groupe de thérapsides majeurs divergent, les cinq autres groupes formant les Eutherapsida. Au sein même de ce clade, les gorgonopsiens sont le groupe frère des euthériodontes, formant ensemble le taxon Theriodontia. Hopson et Barghausen ne sont pas parvenus à une conclusion sur la façon dont les dinocéphales, les anomodontes et les thériodontes seraient liés les uns aux autres, mais des études ultérieures suggèrent que les anomodontes et les thériodontes devraient être classés ensemble au sein des Neotherapsida. Cependant, il reste un débat sur ces relations, en particulier certaines études qui suggèrent que les anomodontes, et non les gorgonopsiens, sont le taxon frère des euthériodontes, d'autres études montrent que les dinocéphales et les anomodontes forment un clade, et la position phylogénétique ainsi que la monophylie des biarmosuchiens restant controversé.
En plus des six grands groupes, il existe des taxons aux classifications incertaines. L'un des exemples les plus connus est celui de Raranimus, datant du début du Permien moyen de l'actuelle Chine, qui est le thérapside le plus basal ainsi que le plus ancien identifié à ce jour[15],[21]. Le genre américain datant du Permien inférieur, Tetraceratops, est depuis longtemps suspecté d'être le thérapside le plus basal connu, étant plus primitif encore que Raranimus[22],[23]. Cependant, une étude de suggère qu'il est plus probable qu'il s'agisse d'un sphénacodonte apparenté plutôt que d'être un vrai représentant des thérapsides[24].
| ||||||||||||||||||||||||||||||||||||||||||
| Cladogramme des thérapsides selon T. S. Kemp ()[25]. |
Galerie

.jpg.webp)
 Lystrosaurus, un anomodonte.
Lystrosaurus, un anomodonte.
 Moschorhinus, un thérocéphale.
Moschorhinus, un thérocéphale.
Notes et références
Notes
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Therapsid » (voir la liste des auteurs).
- Le mot « thérapside » provient du grecs ancien θηρίον / therion signifiant « bête, animal sauvage » et ἁψίς / apsís voulant dire « arc ». L'ensemble pouvant se traduire par « arc de bêtes », en raison de leurs fosses temporales plus grande que celle des autres synapsides.
Références
- (en) R. Broom, « On the use of the term Anomodontia », Records of the Albany Museum, vol. 1, no 4, , p. 266-269.
- (en) Edward D. Cope, « The Theromorphous Reptilia », The American Naturalist, vol. 12, no 12, , p. 829–830 (DOI 10.1086/272251
 )
) - (en) R. L. Carroll, Vertebrate Paleontology and Evolution, New York, W. H. Freeman and Company, , 698 (ISBN 978-0-7167-1822-2)
- (en) Julien Benoit, Fernando Abdala, Paul Manger et Bruce Rubidge, « The sixth sense in mammalians forerunners: variability of the parietal foramen and the evolution of the pineal eye in South African Permo-Triassic eutheriodont therapsids », Acta Palaeontologica Polonica, (ISSN 0567-7920, DOI 10.4202/app.00219.2015 , S2CID 59143925)
- (en) Kévin Rey, Romain Amiot, François Fourel, Fernando Abdala, Frédéric Fluteau, Nour-Eddine Jalil, Jun Liu, Bruce S Rubidge, Roger MH Smith, J Sébastien Steyer, Pia A Viglietti, Xu Wang et Christophe Lécuyer, « Oxygen isotopes suggest elevated thermometabolism within multiple Permo-Triassic therapsid clades », eLife, vol. 6, , –28589 (ISSN 2050-084X, PMID 28716184, PMCID 5515572, DOI 10.7554/eLife.28589 )
- (en) Mathieu G. Faure-Brac et Jorge Cubo, « Were the synapsids primitively endotherms? A palaeohistological approach using phylogenetic eigenvector maps », Philosophical Transactions of the Royal Society B: Biological Sciences, vol. 375, no 1793, , p. 20190138 (ISSN 1471-2970, PMID 31928185, PMCID 7017441, DOI 10.1098/rstb.2019.0138 )
- (en) Elis Newham, Pamela G. Gill, Philippa Brewer, Michael J. Benton, Vincent Fernandez, Neil J. Gostling, David Haberthür, Jukka Jernvall, Tuomas Kankaanpää, Aki Kallonen, Charles Navarro, Alexandra Pacureanu, Kelly Richards, Kate Robson Brown, Philipp Schneider, Heikki Suhonen, Paul Tafforeau, Katherine A. Williams, Berit Zeller-Plumhoff et Ian J. Corfe, « Reptile-like physiology in Early Jurassic stem-mammals », Nature Communications, vol. 11, no 1, , p. 5121 (ISSN 2041-1723, PMID 33046697, PMCID 7550344, DOI 10.1038/s41467-020-18898-4 , Bibcode 2020NatCo..11.5121N)
- (en) James A. Hopson, « The Role of Foraging Mode in the Origin of Therapsids: Implications for the Origin of Mammalian Endothermy », Fieldiana Life and Earth Sciences, vol. 5, , p. 126–148 (ISSN 2158-5520, DOI 10.3158/2158-5520-5.1.126, S2CID 84471370, lire en ligne)
- (en) P. K. Chudinov, « Structure of the integuments of theromorphs », Doklady Akad. Nauk SSSR, vol. 179, , p. 226–229
- (en) Roger M.H. Smith et Jennifer Botha-Brink, « Morphology and composition of bone-bearing coprolites from the Late Permian Beaufort Group, Karoo Basin, South Africa », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 312, nos 1–2, , p. 40–53 (ISSN 0031-0182, DOI 10.1016/j.palaeo.2011.09.006, Bibcode 2011PPP...312...40S, lire en ligne)
- (en) Piotr Bajdek, Martin Qvarnström, Krzysztof Owocki, Tomasz Sulej, Andrey G. Sennikov, Valeriy K. Golubev et Grzegorz Niedźwiedzki., « Microbiota and food residues including possible evidence of pre-mammalian hair in Upper Permian coprolites from Russia. », Lethaia, vol. 49, no 4, , p. 455–477 (DOI 10.1111/let.12156)
- (en) Julien Benoit, Paul R. Manger et Bruce S. Rubidge, « Palaeoneurological clues to the evolution of defining mammalian soft tissue traits », Scientific Reports, vol. 6, no 1, , p. 25604 (PMID 27157809, PMCID 4860582, DOI 10.1038/srep25604 , Bibcode 2016NatSR...625604B).
- (en) Julien Benoit, Irina Ruf, Juri A. Miyamae, Vincent Fernandez, Pablo Gusmão Rodrigues et Bruce S. Rubidge, « The Evolution of the Maxillary Canal in Probainognathia (Cynodontia, Synapsida): Reassessment of the Homology of the Infraorbital Foramen in Mammalian Ancestors », Journal of Mammalian Evolution, vol. 27, no 3, , p. 329–348 (ISSN 1573-7055, DOI 10.1007/s10914-019-09467-8, S2CID 156055693, lire en ligne)
- (en) Richard Estes, « Cranial anatomy of the cynodont reptile Thrinaxodon liorhinus », Bulletin Museum of Comparative Zoology, vol. 125, , p. 165–180
- (en) J. Liu, Rubidge, B et Li, J., « New basal synapsid supports Laurasian origin for therapsids », Acta Palaeontologica Polonica, vol. 54, no 3, , p. 393–400 (DOI 10.4202/app.2008.0071 , lire en ligne [PDF], consulté le )
- (en) Synapsid Classification & Apomorphies
- Adam. K. Huttenlocker et Elizabeth Rega, Forerunners of Mammals: Radiation, Histology, Biology, Indiana University Press, , 90–119 p. (ISBN 978-0253005335, lire en ligne), « Chapter 4. The Paleobiology and Bone Microstructure of Pelycosauriangrade Synapsids »
- (en) Benoit, Norton, Manger et Rubidge, « Reappraisal of the envenoming capacity of Euchambersia mirabilis (Therapsida, Therocephalia) using μCT-scanning techniques », PLoS ONE, vol. 12, no 2, , e0172047 (PMID 28187210, PMCID 5302418, DOI 10.1371/journal.pone.0172047 , Bibcode 2017PLoSO..1272047B)
- (en) A. G. Martinelli, M. B. Soares et C. Schwanke, « Two New Cynodonts (Therapsida) from the Middle-Early Late Triassic of Brazil and Comments on South American Probainognathians », PLOS ONE, vol. 11, no 10, , e0162945 (ISSN 1932-6203, DOI 10.1371/journal.pone.0162945 )
- (en) Kenneth D. Angielczyk et Christian F. Kammerer, Mammalian Evolution, Diversity and Systematics, De Gruyter, , 117–198 p. (ISBN 978-3-11-034155-3, DOI 10.1515/9783110341553-005, S2CID 92370138), « Non-Mammalian synapsids: the deep roots of the mammalian family tree »
- (en) A. Duhamel, J. Benoit, B. S. Rubidge et J. Liu, « A re-assessment of the oldest therapsid Raranimus confirms its status as a basal member of the clade and fills Olson's gap », The Science of Nature, vol. 108, no 4, , p. 26 (ISSN 0028-1042, PMID 34115204, DOI 10.1007/s00114-021-01736-y, Bibcode 2021SciNa.108...26D, S2CID 235403632, lire en ligne)
- (en) M. Laurin et R. R. Reisz, « The osteology and relationships of Tetraceratops insignis, the oldest known therapsid », Journal of Vertebrate Paleontology, vol. 16, no 1, , p. 95–102 (DOI 10.1080/02724634.1996.10011287)
- (en) Eli Amson et Michel Laurin, « On the Affinities of Tetraceratops insignis, an Early Permian Synapsid », Acta Palaeontologica Polonica, vol. 56, no 2, , p. 301–312 (ISSN 0567-7920, DOI 10.4202/app.2010.0063 , S2CID 56425905, lire en ligne)
- (en) Frederik Spindler, « The skull of Tetraceratops insignis (Synapsida, Sphenacodontia) », Palaeovertebrata, vol. 43, no 1, , e1 (DOI 10.18563/pv.43.1.e1 , S2CID 214247325)
- (en) T. S. Kemp, « The origin and early radiation of the therapsid mammal‐like reptiles: a palaeobiological hypothesis », Journal of Evolutionary Biology, vol. 19, no 4, , p. 1231–1247 (PMID 16780524, DOI 10.1111/j.1420-9101.2005.01076.x , S2CID 3184629)
Voir aussi
Articles connexes
Bibliographie
- (en) M. J. Benton, Vertebrate Palaeontology, 3rd Edition, Wiley-Blackwell, (ISBN 978-0-632-05637-8)
- (en) R. L. Carroll, Vertebrate Paleontology & Evolution, New York, W. H. Freeman and Company, (ISBN 978-0716718222)
- (en) T. S. Kemp, The origin and evolution of mammals, Oxford University Press, (ISBN 978-0198507611)
- (en) A. S. Romer, Vertebrate Paleontology 3rd Edition, University of Chicago Press, (ISBN 978-0226724881)
- (en) A. F. Bennett et J. A. Ruben, The ecology and biology of mammal-like reptiles, Washington, D.C., Smithsonian Institution Press, (ISBN 978-0874745245, lire en ligne)
Liens externes
- "Therapsida: Mammals and extinct relatives" sur Tree of Life
- "Therapsida: overview" sur Palaeos (en)
Références taxonomiques
- Ressources relatives au vivant :
- (cs + en) BioLib
- (en) Paleobiology Database
- (en) Interim Register of Marine and Nonmarine Genera
- Notices dans des dictionnaires ou encyclopédies généralistes :
 Portail de la paléontologie
Portail de la paléontologie  Portail des mammifères
Portail des mammifères  Portail origine et évolution du vivant
Portail origine et évolution du vivant