Therocephalia
Thérocéphales • Thérocéphaliens

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Classe | Synapsida |
| Ordre | Therapsida |
| Clade | Eutheriodontia |
Taxons de rang inférieur
- † Crapartinella
- † Gorynychus
- † Theriodesmus
- † Trochosuchidae
- † Lycosuchidae
- † Scylacosauria
Les thérocéphales, parfois appelés thérocéphaliens (Therocephalia), forment un sous-ordre éteint de thérapsides euthériodontes ayant vécu du milieu du Permien jusqu'au milieu du Trias. Le nom du taxon, signifiant littéralement « têtes de bêtes », fait allusion à leur grand crâne et à la structure de leurs dents. Les thérocéphales sont les plus proches parents des cynodontes, le groupe qui donnera plus tard naissance aux mammifères. Cette parenté est illustrée par diverses caractéristiques squelettiques. La phylogénie des thérocéphales est souvent contestée, car la monophylie du groupe ainsi que les relations de ses membres ne sont pas claires.
Les fossiles de thérocéphales sont nombreux dans le supergroupe du Karoo, situé en Afrique du Sud, mais sont également connus de Russie, de Chine, de Tanzanie, de Zambie et même d'Antarctique. Les premiers fossiles de thérocéphales, découverts dans les gisements datant du Permien moyen d'Afrique du Sud, soutiennent une origine du Gondwana pour ces derniers, qui semblent s'être propagés rapidement à travers la planète. Bien que presque toutes les lignées de thérocéphales s'éteignent lors de l'extinction Permien-Trias, quelques représentants du sous-groupe appelé Eutherocephalia, figurent, avec les cynodontes et les dicynodontes (le seul groupe survivant des anomodontes), parmi les rares groupes de synapsides à survivre à cet événement. Certains genres appartenant à ce groupe sont suspectés d'avoir possédé du venin, ce qui en ferait les plus anciens tétrapodes connus à disposer de cette caractéristique. Les derniers thérocéphales s'éteignent au début du Trias moyen, peut-être en raison du changement climatique et de la concurrence avec les cynodontes et de divers groupes de sauropsides, notamment les premiers dinosaures théropodes, qui commencent à apparaitre lors de cette période.
Description
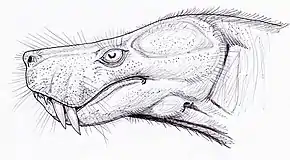
Comme les gorgonopsiens et de nombreux cynodontes, la plupart des thérocéphales seraient vraisemblablement carnivores. Les thérocéphales antérieurs sont, à bien des égards, aussi primitifs que les gorgonopsiens, mais ils présentent certaines caractéristiques avancées. Il y a un élargissement de la fosse temporale pour une plus grande fixation du muscle adducteur de la mâchoire et une réduction des phalanges à la formule mammalienne. La présence d'un palais secondaire naissant chez les thérocéphales avancés est une autre caractéristique partagée avec les cynodontes. La découverte de crêtes maxilloturbinales sous des formes telles que le thérocéphales primitif Glanosuchus (en), suggère qu'au moins certains membres du groupes ont pu avoir le sang chaud[1].

Les thérocéphales ultérieurs comprenent les baurioïdes avancés, qui portent certaines caractéristiques des thériodontes à un degré élevé de spécialisation. Par exemple, les petits baurioïdes et les herbivores Bauria n'ont pas de barre postorbitaire ossifiée séparant l'orbite de la fosse temporale, une condition typique des mammifères primitifs. Ces caractéristiques avancées et d'autres conduisent à l'opinion de longue date, maintenant rejetée, que les trithélodontidés et même certains premiers mammifères serait issus d'une tige de thérocéphales baurioïdes[2]. De telles caractéristiques de mammifères semblent avoir évolué en parallèle parmi un certain nombre de groupes thérapsides différents, même au sein de Therocephalia[1].
Plusieurs modes de vie plus spécialisés sont suggérés pour certains thérocéphales. De nombreuses petites formes, comme les Ictidosuchidés (en), sont interprétées comme des animaux aquatiques. Les preuves de modes de vie aquatiques comprennent des anneaux sclérotiques, qui peuvent stabiliser l'œil sous la pression de l'eau, et des articulations crâniennes fortement développées, qui peuvent soutenir le crâne lors de la consommation de gros poissons et d'invertébrés aquatiques. Certains membres du groupe comme les bauriidés (en) ou les nanictidopidés ont des dents larges avec de nombreuses crêtes similaires à celles des mammifères, et peuvent être des herbivores[3].
De nombreux petits thérocéphales ont de petites fosses sur leurs museau qui supporteraient probablement des vibrisses. En 1994, le paléontologue russe Leonid Tatarinov propose que ces fosses fassent partie d'un système d'électroperception chez les thérocéphales aquatiques[4]. Cependant, il est plus probable que ces fosses soient des versions agrandies de celles censées supporter des moustaches, ou des trous pour les vaisseaux sanguins dans une lèvre charnue[3]. Les genres Euchambersia, Megawhaitsia et Ichibengops, datant du Lopingien, attirent particulièrement l'attention des paléontologues, car les crânes fossiles qui leur sont attribués possèdent des structures qui laissent suggérer que ces deux animaux disposaient des organes de distribution de venin, ce qui en ferait de ces derniers, les plus vieux tétrapodes connus à disposer de tels caractéristiques[5],[6],[7].
Histoire évolutive
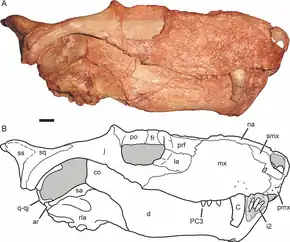
Les thérocéphales évoluent en tant que première lignée de thérapsides euthériodontes pré-mammaliens et sont le groupe frère des cynodontes, qui comprend les mammifères et leurs ancêtres. Les thérocéphales sont au moins aussi anciens qu'une troisième grande branche de thérapsides, les gorgonopsiens, auxquels ils se ressemblent par de nombreux traits primitifs. Par exemple, de nombreux thérocéphales primitifs possèdent de longues canines semblables à celles des gorgonopsiens. Cependant, les gorgonopsiens se sont éteint lors de l'extinction du Permien, les thérocéphales persistant jusqu'au début du Trias moyen.
Alors que l'ascendance commune avec les cynodontes (et, par conséquent, les mammifères) explique de nombreuses similitudes entre ces groupes, certains scientifiques pensent que d'autres similitudes peuvent être mieux attribuées à une évolution convergente, comme la perte de la barre postorbitaire sous certaines formes, une formule phalangienne mammalienne, et une certaine forme de palais secondaire dans la plupart des taxons. Les thérocéphales et les cynodontes survivent de l'extinction Permien-Trias, mais, tandis que les thérocéphales s'éteignent rapidement, les cynodontes connaissent une rapide diversification. Les thérocéphales connaissent une diminution du taux de cladogenèse, ce qui signifie que peu de nouveaux groupes apparaissent après l'extinction. La plupart des lignées de thérocéphales du Trias sont originaires du Permien supérieur et n'existent que sur une courte période[8], s'éteignant à la fin de l'Anisien[9].
Classification
Taxonomie

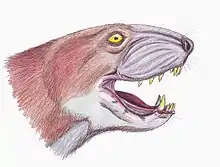
Certains clades de thérocéphales précédemment reconnus se révèlent aujourd'hui obsolète. Par exemple, les Scaloposauridae sont classés sur la base de fossiles avec des caractéristiques principalement juvéniles, représentent probablement des spécimens immatures provenant d'autres familles de thérocéphales connus.
D'autre part, la famille de thérocéphales de la famille des Lycosuchidae, autrefois identifiée par la présence de plusieurs dents caniniformes, est considérée comme représentant d'un groupe non naturel basé sur une étude du remplacement de canine dans ce groupe (van den Heever, ). Cependant, une analyse ultérieure révèle des synapomorphies supplémentaires soutenant la monophylie de ce groupe, et les lycosuchidés sont actuellement considérés comme l'un des taxons le plus basaux au sein des Therocephalia (van den Heever, ).
- Ordre Therapsida
- Sous-ordre Therocephalia
- genre Crapartinella (en)
- genre Gorynychus
- genre Theriodesmus (en)
- Famille Lycosuchidae
- Famille Trochosuchidae (en)
- Clade Scylacosauria
- Sous-ordre Therocephalia
Phylogénie
Cladogramme modifié de Sidor () et Huttenlocker ()[10],[11] :
| ◄ Therapsida |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Vous trouverez ci-dessous un cladogramme modifié à partir d'une analyse publiée par Adam Huttenlocker en [12]. Il est basé sur la matrice de données publiée par Sigurdsen et al. ()[13], qui est lui-même une version modifiée de Huttenlocker et al. (2011)[8]. Six caractères supplémentaires et 22 nouveaux taxons endogroupes sont ajoutés à la matrice de Sigurdsen et al. (), résultant en une matrice qui comprend 58 thérapsides et taxons hors groupe, dont 49 thérocéphales, qui sont notés sur la base de 135 traits morphologiques. Huttenlocker () utilise ce cladogramme pour construire un super-arbre informel pour évaluer les distributions temporelles et phylogénétiques de la taille corporelle chez les euthériodontes du Permien et du Trias[12] :
| ◄ Therapsida |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Notes et références
Notes
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Therocephalia » (voir la liste des auteurs).
Références
- (en) B.S. Rubidge et Sidor, C.A., « Evolutionary patterns among Permo-Triassic therapsids », Annual Review of Ecology, Evolution, and Systematics, vol. 32, , p. 449–480 (DOI 10.1146/annurev.ecolsys.32.081501.114113, lire en ligne [archive du ])
- Fernando Abdala, Bruce S. Rubidge et Juri van den Heever, « The oldest therocephalians (Therapsida, Eutheriodontia) and the early diversification of Therapsida », Palaeontology, vol. 51, no 4, , p. 1011–1024 (ISSN 1475-4983, DOI 10.1111/j.1475-4983.2008.00784.x
 , S2CID 129791548, lire en ligne)
, S2CID 129791548, lire en ligne) - (en) M.F. Ivakhnenko, « Permian and Triassic therocephals (Eutherapsida) of Eastern Europe », Paleontological Journal, vol. 45, no 9, , p. 981–1144 (DOI 10.1134/S0031030111090012)
- (en) L.P. Tatarinov, « On the preservation of rudimentary rostral tubular complex of crossopterygians in theriodonts and on possible development of the electroreceptor systems in some members of this group », Doklady Akademii Nauk, vol. 338, no 2, , p. 278–281
- (en) Ivakhnenko, M.F., « The first Whaitsiid (Therocephalia, Theromorpha) from the Terminal Permian of Eastern Europe », Paleontological Journal, vol. 42, no 4, , p. 409–413 (DOI 10.1134/S0031030108040102, S2CID 140547244)
- (en) Field Museum, « Prehistoric carnivore dubbed 'scarface' discovered in Zambia », Science Daily,
- (en) Benoit, Norton, Manger et Rubidge, « Reappraisal of the envenoming capacity of Euchambersia mirabilis (Therapsida, Therocephalia) using μCT-scanning techniques », PLoS ONE, vol. 12, no 2, , e0172047 (PMID 28187210, PMCID 5302418, DOI 10.1371/journal.pone.0172047 , Bibcode 2017PLoSO..1272047B)
- (en) A.K. Huttenlocker, Sidor, C.A. et Smith, R.M.H., « A new specimen of Promoschorhynchus (Therapsida: Therocephalia: Akidnognathidae) from the Lower Triassic of South Africa and its implications for theriodont survivorship across the Permo-Triassic boundary », Journal of Vertebrate Paleontology, vol. 31, no 2, , p. 405–421 (DOI 10.1080/02724634.2011.546720, S2CID 129242450)
- (en) Henrik Richard Grunert, Brocklehurst, Neil et Fröbisch, Jörg, « Diversity and Disparity of Therocephalia: Macroevolutionary Patterns through Two Mass Extinctions », Scientific Reports, vol. 9, no 5063, , p. 5063 (PMID 30911058, PMCID 6433905, DOI 10.1038/s41598-019-41628-w, Bibcode 2019NatSR...9.5063G)
- (en) C.A. Sidor, « Simplification as a trend in synapsid cranial evolution », Evolution, vol. 55, no 7, , p. 1419–1442 (PMID 11525465, DOI 10.1554/0014-3820(2001)055[1419:saatis]2.0.co;2)
- (en) A. Huttenlocker, « An investigation into the cladistic relationships and monophyly of therocephalian therapsids (Amniota: Synapsida) », Zoological Journal of the Linnean Society, vol. 157, no 4, , p. 865–891 (DOI 10.1111/j.1096-3642.2009.00538.x )
- (en) A. K. Huttenlocker, « Body Size Reductions in Nonmammalian Eutheriodont Therapsids (Synapsida) during the End-Permian Mass Extinction », PLOS ONE, vol. 9, no 2, , e87553 (PMID 24498335, PMCID 3911975, DOI 10.1371/journal.pone.0087553 , Bibcode 2014PLoSO...987553H)
- (en) T. Sigurdsen, A. K. Huttenlocker, S. P. Modesto, T. B. Rowe et R. Damiani, « Reassessment of the morphology and paleobiology of the therocephalian Tetracynodon darti (Therapsida), and the phylogenetic relationships of Baurioidea » (Submitted manuscript), Journal of Vertebrate Paleontology, vol. 32, no 5, , p. 1113 (DOI 10.1080/02724634.2012.688693, S2CID 84457790)
Voir aussi
Articles connexes
Liens externes
- (en) Référence Tree of Life Web Project : Therocephalia
- (en) Référence Paleobiology Database : Therocephalia Broom 1903
- Theriodontia - sur Paleos
 Portail de la paléontologie
Portail de la paléontologie