Thalamocortical radiations
| Thalamocortical radiations | |
|---|---|
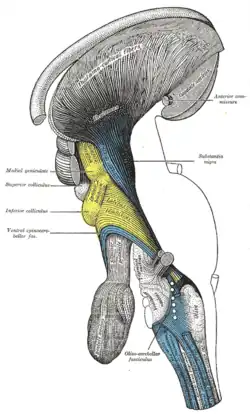 Deep dissection of brain-stem. Lateral view. (Thalamocortical fibers labeled at center top.) | |
 Tractography of thalamocortical radiation | |
| Details | |
| From | Thalamus |
| To | Cortex |
| Identifiers | |
| Latin | Radiatio thalamocorticalis, tractus thalamocorticalis |
| NeuroNames | 1337 |
| Anatomical terms of neuroanatomy | |
Thalamocortical radiations are the fibers between the thalamus and the cerebral cortex.[1]
Structure
Thalamocortical (TC) fibers have been referred to as one of the two constituents of the isothalamus, the other being micro neurons. Thalamocortical fibers have a bush or tree-like appearance as they extend into the internal capsule and project to the layers of the cortex. The main thalamocortical fibers extend from different nuclei of the thalamus and project to the visual cortex, somatosensory (and associated sensori-motor) cortex, and the auditory cortex in the brain. Thalamocortical radiations also innervate gustatory and olfactory pathways, as well as pre-frontal motor areas. Visual input from the optic tract is processed by the lateral geniculate nucleus of the thalamus, auditory input in the medial geniculate nucleus, and somatosensory input in the ventral posterior nucleus of the thalamus. Thalamic nuclei project to cortical areas of distinct architectural organization and relay the processed information back to the area of original activity in the thalamus via corticothalamic (CT) fibers.[2] The thalamic reticular nucleus (TRN) receives incoming signals via corticothalamic pathways and regulates activity within the thalamus accordingly.[3] Cortico-thalamic feedback neurons are mostly found in layer VI of the cortex.[4] Reciprocal CT projections to the thalamus are of a higher order than, and synapse with, the TRN in much greater number than do thalamocortical projections to cortex.[5] This suggests that the cortex has a much bigger role in top down processing and regulation of thalamic activity than do the processes originating in thalamic interneurons. Large-scale frequency oscillations and electrical rhythms have also been shown to regulate TC activity for long periods of time, as is evident during the sleep cycle.[6] Other evidence suggests CT modulation of TC rhythms can occur over different time scales, adding even more complexity to their function.[4]
Relay cells
Thalamic interneurons process sensory information and signal different regions of the thalamic nuclei. These nuclei extend to relay cells, which in turn innervate distinct areas of the cortex via thalamocortical fibers. Either specifically or nonspecifically, TC relay cells project specifically to organized areas of the cortex directly and nonspecifically project to large areas of cortex through the innervation of many interconnected collateral axons.[7] According to Jones (2001) there are two primary types of relay neurons in the thalamus of primates–core cells and matrix cells–each creating distinct pathways to various parts and layers throughout the cerebral cortex.[5] Matrix cells of the thalamus, or calbindin-immuno-reactive neurons (CIR neurons), are widely distributed and diffusely dispersed in each of the nuclei of the dorsal thalamus. In comparison, parvalbumin immuno-reactive neurons (PIR neurons) can be found only in principal sensory and motor relay nuclei, and in the pulvinar nuclei as well as the intralaminar nuclei. The PIR neurons cluster together creating "densely terminating afferent fibers…forming a core imposed on a diffuse background matrix of PIR cells" (Jones 2001). PIR cells tend to project upon the cerebral cortex and terminate in an organized topographic manner in specifically localized zones (in deep layer III and in the middle layer IV). In contrast, CIR cells have dispersed projections wherein various adjacent cells connect to non-specific different cortical areas. CIR axons seem to terminate primarily in the superficial layers of the cortex: layers I, II, and upper III.[5]
Function
Thalamocortical signaling is primarily excitatory, causing the activation of corresponding areas of the cortex, but is mainly regulated by inhibitory mechanisms. The specific excitatory signaling is based upon glutamatergic signaling, and is dependent on the nature of the sensory information being processed. Recurrent oscillations in thalamocortical circuits also provide large-scale regulatory feedback inputs to the thalamus via GABAergic neurons that synapse in the TRN.[8] In a study done by Gibbs, Zhang, Shumate, and Coulter (1998) it was found that endogenously released zinc blocked GABA responses within the TC system specifically by interrupting communication between the thalamus and the connected TRN.[8] Computational neuroscientists are particularly interested in thalamocortical circuits because they represent a structure that is disproportionally larger and more complex in humans than other mammals (when body size is taken into account), which may contribute to humans' special cognitive abilities.[9] Evidence from one study (Arcelli et al. 1996) offers partial support to this claim by suggesting that thalamic GABAergic local circuit neurons in mammalian brains relate more to processing ability compared to sensorimotor ability, as they reflect an increasing complexity of local information processing in the thalamus.[7] It is proposed that core relay cells and matrix cells projecting from the dorsal thalamus allow for synchronization of cortical and thalamic cells during "high-frequency oscillations that underlie discrete conscious events",[5] though this is a heavily debated area of research.
Projections
The majority of thalamocortical fibers project to layer IV of the cortex, wherein sensory information is directed to other layers where they either terminate or connect with axons collaterally depending on type of projection and type of initial activation.[10] Activation of the thalamocortical neurons relies heavily on the direct and indirect effects of Glutamate, which causes EPSP's at terminal branches in the primary sensory cortices.[11]
Somatosensory areas
Primarily, thalamocortical somatosensory radiation from the VPL, VPM and LP nuclei extends to the primary and secondary somatosensory areas, terminating in cortical layers of the lateral postcentral gyrus. S1 receives parallel thalamocortical radiations from the posterior medial nucleus and the VPN.[12] Projections from the VPN to the postcentral gyrus account for the transfer of sensory information concerning touch and pain. Several studies indicate that parallel innervations to S1 and also S2 via thalamocortical pathways result in the processing of nociceptive and non-nociceptive information. Non-specific projections to sensori-motor areas of the cortex may in part have to do with the relationship between non-noci-receptive processing and motor functions.[13] Past research shows a link between S1 and M1, creating a thalamocortical sensori-motor circuit. When this circuit becomes disrupted symptoms are produced similar to those that accompany Multiple sclerosis, suggesting thalamocortical rhythms are involved in regulating sensori-motor pathways in a highly specialized manner.[14] TC-CT rhythms evident during sleep act to inhibit these thalamocortical fibers so as to maintain the tonic cycling of low frequency waves and the subsequent suppression of motor activity.[6]
Visual areas
The Lateral geniculate and pulvinar nuclei project to and terminate in V1, and carry motor information from the brain stem as well as other sensory input from the optic tract. The visual cortex connects with other sensory areas which allows for the integration of cognitive tasks such as selective and directed attention, and pre-motor planning, in relation to the processing of incoming visual stimuli.[15] Models of the pulvinar projections to the visual cortex have been proposed by several imaging studies, though the mapping of pulvinar projections has been a difficult task due to the fact that pulvinar subdivisions are not conventionally organized and have been difficult to visualize using structural MRI.[15] Evidence from several studies supports the idea that the Pulvinar nuclei and superior colliculus receive descending projections from CT fibers while TC fibers extending from the LGN carry visual information to the various areas of the visual cortex near the calcarine fissure.
Auditory areas
Thalamocortical axons project primarily from the medial geniculate nucleus via the sublenticular region of the internal capsule, and terminate in an organized topographic manner in the transverse temporal gyri.[16] MMGN radiations terminate in specific locations while thalamocortical fibers from the VMGN terminate in nonspecific clusters of cells and form collateral connections to neighboring cells.[17] Research done by staining the brains of macaque monkeys reveals projections from the ventral nucleus mainly terminating in layers IV and IIIB, with some nonspecific clusters of PIR cells terminating in layers I, II, IIIA, and VI. Fibers from the dorsal nuclei were found to project more directly to the primary auditory area, with most axons terminating in layer IIIB. The magnocellular nucleus projected a small amount of PIR cells with axons mainly terminating in layer 1, though large regions of the middle cortical layers were innervated through collaterally connected CIR neurons.[18] Past research suggests that the thalamocortical-auditory pathway may be the only neural correlate that can explain a direct translation of frequency information to the cortex via specific pathways.[19]
Motor areas
The primary motor cortex receives terminating thalamocortical fibers from the VL nucleus of the thalamus. This is the primary pathway involved in the transference of cerebellar input to the primary motor cortex. The VA projects widely across the inferior parietal and premotor cortex. Other Non-specific thalamocortical projections, those that originate in the dorsal-medial nuclei of the thalamus, terminate in the prefrontal cortex and have subsequent projections to associative premotor areas via collateral connections.[20] The Cortico-basal ganglia-thalamocortical loop has been traditionally associated with reward-learning and though has also been noted by some researchers to have a modulatory effect on thalamocortical network functioning–this is due to inherent activation of the premotor areas connecting the VA nucleus with the cortex.[20]
Clinical significance
Absence seizures
Thalamocortical radiations have been researched extensively in the past due to their relationship with attention, wakefulness, and arousal. Past research has shown how an increase in spike-and-wave activity within the TC network can disrupt normal rhythms involved with the sleep-wakefulness cycle, ultimately causing absence seizures and other forms of epileptic behavior. Burst firing within a part of the TC network stimulates GABA receptors within the thalamus causing moments of increased inhibition, leading to frequency spikes, which offset oscillation patterns.[21][22] Another study done on rats suggests during spike-and-wave seizures, thalamic rhythms are mediated by local thalamic connections, while the cortex controls the synchronization of these rhythms over extended periods of time. Thalamocortical dysrhythmia is a term associated with spontaneously reoccurring low frequency spike-and-wave activity in the thalamus, which causes symptoms normally associated with impulse control disorders such as obsessive compulsive disorder, Parkinson's disease, attention deficit hyperactivity disorder, and other forms of chronic psychosis.[23] Other evidence has shown how reductions in the distribution of connections of nonspecific thalamocortical systems is heavily associated with loss of consciousness, as can be seen with individuals in a vegetative state, or coma.[24]
Prefrontal lobotomy
The bilateral interruption or severing of the connection between thalamocortical radiations the medial and anterior thalamic nuclei results in a prefrontal lobotomy, which causes a drastic personality change and a subdued behavioral disposition without cortical injury.[25]
Research
Evolutionary theories of consciousness
Theories of consciousness have been linked to thalamocortical rhythm oscillations in TC-CT pathway activity. One such theory, the dynamic core theory of conscious experience, proposes four main pillars in support of conscious awareness as a consequence of dorsal thalamic activity:[26]
- the results of cortical computations underlay consciousness
- vegetative states and general anesthetics work primarily to disrupt normal thalamic functioning
- the anatomy and physiology of the thalamus implies consciousness
- neural synchronization accounts for the neural basis of consciousness.
This area of research is still developing, and most current theories are either partial or incomplete.
References
- ↑ University of Washington (1991). "Thalamocortical radiations". washington.edu.
- ↑ Granger H.R.; Hearn R.A. (2007). "Models of thalamocortical system". Scholarpedia the peer-reviewed open-access encyclopedia. Vol. 2. p. 1796. Bibcode:2007SchpJ...2.1796G. doi:10.4249/scholarpedia.1796.
- ↑ Jones E.G. (1975). "Some aspects of the organization of the thalamic reticular complex". Journal of Computational Neurology. 162 (3): 285–308. doi:10.1002/cne.901620302. PMID 1150923. S2CID 28724898.
- 1 2 Briggs F.; Usrey W.M. (2008). "emerging views of corticothalamic function". Current Opinion in Neurobiology. 18 (4): 403–407. doi:10.1016/j.conb.2008.09.002. PMC 2626162. PMID 18805486.
- 1 2 3 4 Jones E.G. (2002). "Thalamic circuitry and thalamocortical synchrony". Philosophical Transactions of the Royal Society B. 357 (1428): 1659–1673. doi:10.1098/rstb.2002.1168. PMC 1693077. PMID 12626002.
- 1 2 Hill S.; Tononi G. (2005). "modeling sleep and wakefulness in the thalamocortical system". Journal of Neurophysiology. 93 (3): 1671–1698. doi:10.1152/jn.00915.2004. PMID 15537811.
- 1 2 Arcelli P.; Frassoni C.; Regondi M.C.; Biasi S.D.; Spreafico R. (1997). "GABAergic neurons in mammalian thalamus: a marker of thalamic complexity?". Brain Research Bulletin. 42 (1): 27–37. doi:10.1016/S0361-9230(96)00107-4. PMID 8978932. S2CID 23211449.
- 1 2 Gibbs J.W.; Zhang Y.; Shumate M.D.; Coulter D.A. (1999). "Regionally selective blockade of GABAergic inhibition by zinc in the thalamocortical system: functional significance". Journal of Neurophysiology. 83 (3): 1510–1521. doi:10.1152/jn.2000.83.3.1510. PMID 10712476.
- ↑ Rodriguez A.W.; Whitson J.; Granger R. (2004). "Derivation and analysis of basic computational operations of thalamocortical circuits". Journal of Cognitive Neuroscience. 16 (5): 856–877. CiteSeerX 10.1.1.132.8042. doi:10.1162/089892904970690. PMID 15200713. S2CID 5652129.
- ↑ Palmer S.L.; Noctor S.C.; Jablonska B.; Juliano S.L. (2001). "Laminar specific alterations of thalamocortical projections in organotypic cultures following layer 4 disruption in ferret somatosensory cortex". European Journal of Neuroscience. 13 (8): 1559–71. doi:10.1046/j.0953-816x.2001.01519.x. PMID 11328350. S2CID 10933349.
- ↑ Kharazia V.N.; Weinberg R.J. (1994). "Glutamate in thalamic fibers terminating in layer IV of primary sensory cortex". Journal of Neuroscience. 14 (10): 6021–6032. doi:10.1523/JNEUROSCI.14-10-06021.1994. PMC 6577001. PMID 7931559.
- ↑ Viaene A.N.; et al. (2011). "synaptic properties of thalamic input to layers 2/3 and 4 of primary somatosensory and auditory cortices". Journal of Neurophysiology. 105 (1): 279–292. doi:10.1152/jn.00747.2010. PMC 3023380. PMID 21047937.
- ↑ Liang; et al. (2011). "Parallel processing of nociceptive and non-nociceptive somatosensory information in the human primary and secondary somatosensory cortices: Evidence from dynamic causal modeling of functional magnetic resonance imaging data". Journal of Neuroscience. 31 (24): 8976–8985. doi:10.1523/JNEUROSCI.6207-10.2011. PMC 6622932. PMID 21677181.
- ↑ Del'Acqua; et al. (2010). "Thalamocortical sensorimotor circuit in multiple sclerosis: an integrated structural and electrophysiological assessment". Human Brain Mapping. 3 (10): 1588–1600. doi:10.1002/hbm.20961. PMC 6871076. PMID 20162580.
- 1 2 Van Essen D.C. (2005). "corticocortical and thalamocortical information flow in the primate visual system". Cortical Function: A View from the Thalamus. Progress in Brain Research. Vol. 149. pp. 173–185. doi:10.1016/S0079-6123(05)49013-5. ISBN 9780444516794. PMID 16226584.
- ↑ Afifi A.K.; Bergman R.A. (December 1, 1997). Functional neuroanatomy. McGraw-Hill Professional. ISBN 978-0070015890.
- ↑ Theyel; et al. (2010). "specific and nonspecific thalamocortical connectivity in the auditory and somatosensory thalamocortical slices". NeuroReport. 21 (13): 861–864. doi:10.1097/WNR.0b013e32833d7cec. PMC 2945241. PMID 20647961.
- ↑ Jones; et al. (1995). "Patchy and laminar terminations of medial geniculate axons in monkey auditory cortex". Journal of Comparative Neurology. 362 (2): 195–208. doi:10.1002/cne.903620204. PMID 8576433. S2CID 34644952.
- ↑ Xiuping; et al. (2011). "contributions of the thalamocortical system towards sound specific auditory plasticity". Neuroscience & Biobehavioral Reviews. 35 (10): 2155–2161. doi:10.1016/j.neubiorev.2011.02.010. PMID 21349286. S2CID 42437671.
- 1 2 Arslan O. neuroanatomical basis of clinical neurology.
- ↑ Kostopoulos G.K. (2001). "Involvement of the thalamocortical system in epileptic loss of consciousness". Epilepsia. 42 (3): 13–19. doi:10.1046/j.1528-1157.2001.042suppl.3013.x. PMID 11520316. S2CID 23495744.
- ↑ Whalley K. (2010). "Inhibition: too much of a good thing?". Nature Reviews Neuroscience. 11 (1): 6–7. doi:10.1038/nrn2782. S2CID 33898199.
- ↑ Llinas; et al. (2003). "Neuropsychiatric thalamocortical dysrhythmia: surgical implications". Thalamus and Related Systems. 2 (2): 103–113. doi:10.1017/S1472928803000104. PMID 12856492.
- ↑ Zhou; et al. (2011). "specific and nonspecific thalamocortical functional connectivity in normal and vegetative states". Consciousness and Cognition. 20 (2): 257–268. doi:10.1016/j.concog.2010.08.003. PMC 3056940. PMID 21078562.
- ↑ Freeman W.; Watts J.K. (1942). "Prefrontal lobotomy; the surgical relief of mental pain". Bulletin of the New York Academy of Medicine. 18 (12): 794–812. PMC 1933933. PMID 19312295.
- ↑ Ward L. M. (2011). "the thalamic dynamic core theory of conscious experience". Consciousness and Cognition. 20 (2): 464–486. doi:10.1016/j.concog.2011.01.007. PMID 21349742. S2CID 6099949.