CD154
CD154, also called CD40 ligand or CD40L, is a protein that is primarily expressed on activated T cells[5] and is a member of the TNF superfamily of molecules. It binds to CD40 on antigen-presenting cells (APC), which leads to many effects depending on the target cell type. In total CD40L has three binding partners: CD40, α5β1 integrin and integrin αIIbβ3. CD154 acts as a costimulatory molecule and is particularly important on a subset of T cells called T follicular helper cells (TFH cells).[6] On TFH cells, CD154 promotes B cell maturation and function by engaging CD40 on the B cell surface and therefore facilitating cell-cell communication.[7] A defect in this gene results in an inability to undergo immunoglobulin class switching and is associated with hyper IgM syndrome.[8] Absence of CD154 also stops the formation of germinal centers and therefore prohibiting antibody affinity maturation, an important process in the adaptive immune system.

History
In 1991, three groups reported discovering CD154. Seth Lederman, Michael Yellin, and Leonard Chess at Columbia University generated a murine monoclonal antibody, 5c8, that inhibited contact-dependent T cell helper function in human cells and which characterized a 32 kDa surface protein transiently expressed on activated CD4+ T cells.[9] Richard Armitage at Immunex cloned a cDNA encoding CD154 by screening an expression library with CD40-Ig.[10] Randolph Noelle at Dartmouth Medical School generated an antibody that bound a 39 kDa protein on murine T cells and inhibited helper function.[11] Noelle contested Lederman's patent, but the challenge (called an interference) was rejected on all counts [12]
Expression
CD40 ligand (CD154) is primarily expressed on activated CD4+ T lymphocytes but is also found in a soluble form. While CD40L was originally described on T lymphocytes, its expression has since been found on a wide variety of cells, including platelets, mast cells, macrophages, basophils, NK cells, B lymphocytes, as well as non-haematopoietic cells (smooth muscle cells, endothelial cells, and epithelial cells).[13]
Specific effects on cells
CD40L plays a central role in costimulation and regulation of the immune response via T cell priming and activation of CD40-expressing immune cells.[14] At least 46 disease-causing mutations in this gene have been discovered.[15]
Macrophages
In the macrophage, the primary signal for activation is IFN-γ from Th1 type CD4 T cells. The secondary signal is CD40L on the T cell, which binds CD40 on the macrophage cell surface. As a result, the macrophage expresses more CD40 and TNF receptors on its surface, which helps increase the level of activation. The activated macrophage can then destroy phagocytosed bacteria and produce more cytokines.
B cells

B cells can present antigens to a specialized group of helper T cells called TFH cells. If an activated TFH cell recognizes the peptide presented by the B cell, the CD40L on the T cell binds to the B cell's CD40, causing B cell activation.[16] The T cell also produces IL-4, which directly influences B cells. As a result of this stimulation, the B cell can undergo rapid cellular division to form a germinal center where antibody isotype switching and affinity maturation occurs, as well as their differentiation to plasma cells and memory B cells. The end-result is a B cell that is able to mass-produce specific antibodies against an antigenic target. Early evidence for these effects were that in CD40 or CD154 deficient mice, there is little class switching or germinal centre formation, and immune responses are severely inhibited.[17]
Endothelial cells
Activation of endothelial cells by CD40L (e.g. from activated platelets) leads to reactive oxygen species production, as well as chemokine and cytokine production, and expression of adhesion molecules such as E-selectin, ICAM-1, and VCAM-1. This inflammatory reaction in endothelial cells promotes recruitment of leukocytes to lesions and may potentially promote atherogenesis.[18] CD40L has shown to be a potential biomarker for atherosclerotic instability.[19]
References
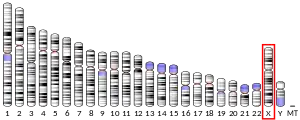
- GRCh38: Ensembl release 89: ENSG00000102245 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000031132 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Lederman S, Yellin MJ, Krichevsky A, Belko J, Lee JJ, Chess L (April 1992). "Identification of a novel surface protein on activated CD4+ T cells that induces contact-dependent B cell differentiation (help)". The Journal of Experimental Medicine. 175 (4): 1091–101. doi:10.1084/jem.175.4.1091. PMC 2119166. PMID 1348081.
- Lederman S, Yellin MJ, Inghirami G, Lee JJ, Knowles DM, Chess L (December 1992). "Molecular interactions mediating T-B lymphocyte collaboration in human lymphoid follicles. Roles of T cell-B-cell-activating molecule (5c8 antigen) and CD40 in contact-dependent help". Journal of Immunology. 149 (12): 3817–26. PMID 1281189.
- Lederman S, Yellin MJ, Cleary AM, Pernis A, Inghirami G, Cohn LE, Covey LR, Lee JJ, Rothman P, Chess L (March 1994). "T-BAM/CD40-L on helper T lymphocytes augments lymphokine-induced B cell Ig isotype switch recombination and rescues B cells from programmed cell death". Journal of Immunology. 152 (5): 2163–71. PMID 7907632.
- "Entrez Gene: CD40LG CD40 ligand (TNF superfamily, member 5, hyper-IgM syndrome)".
- Lederman S, Yellin MJ, Krichevsky A, Belko J, Lee JJ, Chess L (April 1992). "Identification of a novel surface protein on activated CD4+ T cells that induces contact-dependent B cell differentiation (help)". The Journal of Experimental Medicine. 175 (4): 1091–101. doi:10.1084/jem.175.4.1091. PMC 2119166. PMID 1348081.
- Armitage RJ, Fanslow WC, Strockbine L, Sato TA, Clifford KN, Macduff BM, Anderson DM, Gimpel SD, Davis-Smith T, Maliszewski CR, et al. (May 1992). "Molecular and biological characterization of a murine ligand for CD40". Nature. 357 (6373): 80–2. Bibcode:1992Natur.357...80A. doi:10.1038/357080a0. PMID 1374165. S2CID 4336943.
- Noelle RJ, Roy M, Shepherd DM, Stamenkovic I, Ledbetter JA, Aruffo A (July 1992). "A 39-kDa protein on activated helper T cells binds CD40 and transduces the signal for cognate activation of B cells". Proc Natl Acad Sci U S A. 89 (14): 6550–4. Bibcode:1992PNAS...89.6550N. doi:10.1073/pnas.89.14.6550. PMC 49539. PMID 1378631.
- "Patent 5,474,771" (PDF). UNITED STATES PATENT AND TRADEMARK OFFICE. U.S. Patent Office. Archived from the original (PDF) on 2016-03-04. Retrieved 2016-06-18.
- Schönbeck U, Libby P (January 2001). "The CD40/CD154 receptor/ligand dyad". Cellular and Molecular Life Sciences. 58 (1): 4–43. doi:10.1007/PL00000776. PMID 11229815. S2CID 33085593.
- Grewal, IS; Flavell, RA (1998). "CD40 and CD154 in cell-mediated immunity". Annual Review of Immunology. 16: 111–35. doi:10.1146/annurev.immunol.16.1.111. PMID 9597126.
- Šimčíková D, Heneberg P (December 2019). "Refinement of evolutionary medicine predictions based on clinical evidence for the manifestations of Mendelian diseases". Scientific Reports. 9 (1): 18577. Bibcode:2019NatSR...918577S. doi:10.1038/s41598-019-54976-4. PMC 6901466. PMID 31819097.
- Cleary AM, Fortune SM, Yellin MJ, Chess L, Lederman S (October 1995). "Opposing roles of CD95 (Fas/APO-1) and CD40 in the death and rescue of human low density tonsillar B cells". Journal of Immunology. 155 (7): 3329–37. PMID 7561026.
- Grewal, IS; Xu, J; Flavell, RA (7 December 1995). "Impairment of antigen-specific T-cell priming in mice lacking CD40 ligand". Nature. 378 (6557): 617–20. Bibcode:1995Natur.378..617G. doi:10.1038/378617a0. PMID 8524395. S2CID 4259617.
- Szmitko PE, Wang CH, Weisel RD, de Almeida JR, Anderson TJ, Verma S (October 2003). "New markers of inflammation and endothelial cell activation: Part I". Circulation. 108 (16): 1917–23. doi:10.1161/01.CIR.0000089190.95415.9F. PMID 14568885.
- Wang JH, Zhang YW, Zhang P, Deng BQ, Ding S, Wang ZK, Wu T, Wang J (September 2013). "CD40 ligand as a potential biomarker for atherosclerotic instability". Neurological Research. 35 (7): 693–700. doi:10.1179/1743132813Y.0000000190. PMC 3770830. PMID 23561892.
- Lineberry NB, Su LL, Lin JT, Coffey GP, Seroogy CM, Fathman CG (August 2008). "Cutting edge: The transmembrane E3 ligase GRAIL ubiquitinates the costimulatory molecule CD40 ligand during the induction of T cell anergy". Journal of Immunology. 181 (3): 1622–6. doi:10.4049/jimmunol.181.3.1622. PMC 2853377. PMID 18641297.
Further reading
- Parham, Peter (2004). The Immune System (2nd ed.). Garland Science. pp. 169–173. ISBN 0-8153-4093-1.
- Tong AW, Stone MJ (March 1996). "CD40 and the effect of anti-CD40-binding on human multiple myeloma clonogenicity". Leukemia & Lymphoma. 21 (1–2): 1–8. doi:10.3109/10428199609067572. PMID 8907262.
- van Kooten C, Banchereau J (January 2000). "CD40-CD40 ligand". Journal of Leukocyte Biology. 67 (1): 2–17. doi:10.1002/jlb.67.1.2. PMID 10647992. S2CID 35592719.
- Schattner EJ (May 2000). "CD40 ligand in CLL pathogenesis and therapy". Leukemia & Lymphoma. 37 (5–6): 461–72. doi:10.3109/10428190009058499. PMID 11042507. S2CID 39398949.
- Bhushan A, Covey LR (2002). "CD40:CD40L interactions in X-linked and non-X-linked hyper-IgM syndromes". Immunologic Research. 24 (3): 311–24. doi:10.1385/IR:24:3:311. PMID 11817328. S2CID 19537892.
- Cheng G, Schoenberger SP (2002). "CD40 signaling and autoimmunity". Signal Transduction Pathways in Autoimmunity. Current Directions in Autoimmunity. Vol. 5. pp. 51–61. doi:10.1159/000060547. ISBN 3-8055-7308-1. PMID 11826760.
- Subauste CS (February 2002). "CD154 and type-1 cytokine response: from hyper IgM syndrome to human immunodeficiency virus infection". The Journal of Infectious Diseases. 185 (Suppl 1): S83-9. doi:10.1086/338003. PMID 11865444.
- Kornbluth RS (October 2002). "An expanding role for CD40L and other tumor necrosis factor superfamily ligands in HIV infection". Journal of Hematotherapy & Stem Cell Research. 11 (5): 787–801. doi:10.1089/152581602760404595. PMID 12427285.
- Xu Y, Song G (2005). "The role of CD40-CD154 interaction in cell immunoregulation". Journal of Biomedical Science. 11 (4): 426–38. doi:10.1159/000077892. PMID 15153777. S2CID 202658036.
- Law, CL; Grewal, IS (2009). "Therapeutic Interventions Targeting CD40L (CD154) and CD40: The Opportunities and Challenges". Therapeutic Targets of the TNF Superfamily. Advances in Experimental Medicine and Biology. Vol. 647. pp. 8–36. doi:10.1007/978-0-387-89520-8_2. ISBN 978-0-387-89519-2. PMID 19760064.
External links
- CD154+Antigen at the U.S. National Library of Medicine Medical Subject Headings (MeSH)
- Human CD40LG genome location and CD40LG gene details page in the UCSC Genome Browser.
- GeneReviews/NCBI/NIH/UW entry on X-Linked Hyper IgM Syndrome or Immunodeficiency with Hyper-IgM, Type 1
PDB gallery | |
|---|---|
|