Superoxide dismutase
Superoxide dismutase (SOD, EC 1.15.1.1) is an enzyme that alternately catalyzes the dismutation (or partitioning) of the superoxide (O−
2) radical into ordinary molecular oxygen (O2) and hydrogen peroxide (H
2O
2). Superoxide is produced as a by-product of oxygen metabolism and, if not regulated, causes many types of cell damage.[2] Hydrogen peroxide is also damaging and is degraded by other enzymes such as catalase. Thus, SOD is an important antioxidant defense in nearly all living cells exposed to oxygen. One exception is Lactobacillus plantarum and related lactobacilli, which use a different mechanism to prevent damage from reactive O−
2.
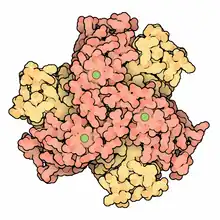
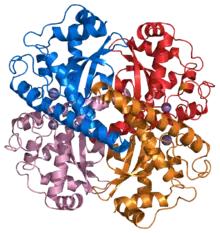 Structure of a human Mn superoxide dismutase 2 tetramer[1] | |||||||||
| Identifiers | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| EC no. | 1.15.1.1 | ||||||||
| CAS no. | 9054-89-1 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| Gene Ontology | AmiGO / QuickGO | ||||||||
| |||||||||
Chemical reaction
SODs catalyze the disproportionation of superoxide:
- 2H+
+ 2O−
2 → O
2 + H
2O
2
In this way, O−
2 is converted into two less damaging species.
The general form, applicable to all the different metal−coordinated forms of SOD, can be written as follows:
- M
(n+1)+−SOD + O−
2 → M
n+−SOD + O
2 - M
n+−SOD + O−
2 + 2H+
→ M
(n+1)+−SOD + H
2O
2
The reactions by which SOD−catalyzed dismutation of superoxide for Cu,Zn SOD can be written as follows:
- Cu2+
−SOD + O−
2 → Cu+
−SOD + O
2 (reduction of copper; oxidation of superoxide) - Cu+
−SOD + O−
2 + 2H+
→ Cu2+
−SOD + H
2O
2 (oxidation of copper; reduction of superoxide)
where M = Cu (n=1); Mn (n=2); Fe (n=2); Ni (n=2) only in prokaryotes.
In a series of such reactions, the oxidation state and the charge of the metal cation oscillates between n and n+1: +1 and +2 for Cu, or +2 and +3 for the other metals .
Types
General
Irwin Fridovich and Joe McCord at Duke University discovered the enzymatic activity of superoxide dismutase in 1968.[3] SODs were previously known as a group of metalloproteins with unknown function; for example, CuZnSOD was known as erythrocuprein (or hemocuprein, or cytocuprein) or as the veterinary anti-inflammatory drug "Orgotein".[4] Likewise, Brewer (1967) identified a protein that later became known as superoxide dismutase as an indophenol oxidase by protein analysis of starch gels using the phenazine-tetrazolium technique.[5]
There are three major families of superoxide dismutase, depending on the protein fold and the metal cofactor: the Cu/Zn type (which binds both copper and zinc), Fe and Mn types (which bind either iron or manganese), and the Ni type (which binds nickel).
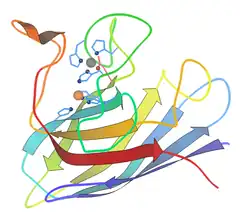
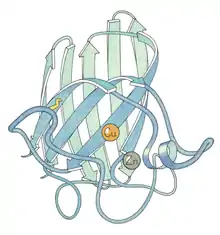 Ribbon diagram of bovine Cu-Zn SOD subunit[6] |
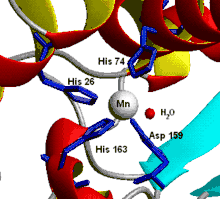
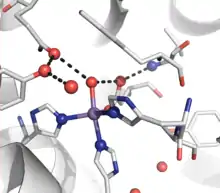 Active site of Human Manganese SOD, manganese shown in purple[7] |
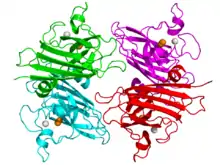
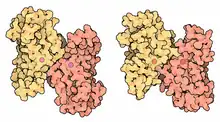 Mn-SOD vs Fe-SOD dimers |
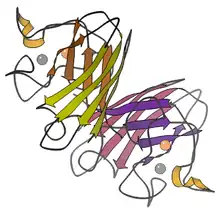
- Copper and zinc – most commonly used by eukaryotes, including humans. The cytosols of virtually all eukaryotic cells contain an SOD enzyme with copper and zinc (Cu-Zn-SOD). For example, Cu-Zn-SOD available commercially is normally purified from bovine red blood cells. The bovine Cu-Zn enzyme is a homodimer of molecular weight 32,500. It was the first SOD whose atomic-detail crystal structure was solved, in 1975.[8] It is an 8-stranded "Greek key" beta-barrel, with the active site held between the barrel and two surface loops. The two subunits are tightly joined back-to-back, mostly by hydrophobic and some electrostatic interactions. The ligands of the copper and zinc are six histidine and one aspartate side-chains; one histidine is bound between the two metals.[9]
- Iron or manganese – used by prokaryotes and protists, and in mitochondria and chloroplasts
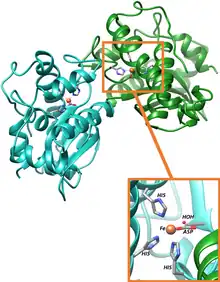 Active site for iron superoxide dismutase
Active site for iron superoxide dismutase- Iron – Many bacteria contain a form of the enzyme with iron (Fe-SOD); some bacteria contain Fe-SOD, others Mn-SOD, and some (such as E. coli) contain both. Fe-SOD can also be found in the chloroplasts of plants. The 3D structures of the homologous Mn and Fe superoxide dismutases have the same arrangement of alpha-helices, and their active sites contain the same type and arrangement of amino acid side-chains. They are usually dimers, but occasionally tetramers.
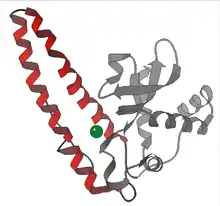
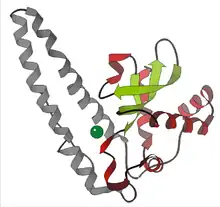
- Manganese – Nearly all mitochondria, and many bacteria, contain a form with manganese (Mn-SOD): For example, the Mn-SOD found in human mitochondria. The ligands of the manganese ions are 3 histidine side-chains, an aspartate side-chain and a water molecule or hydroxy ligand, depending on the Mn oxidation state (respectively II and III).[10]
- Nickel – prokaryotic. This has a hexameric (6-copy) structure built from right-handed 4-helix bundles, each containing N-terminal hooks that chelate a Ni ion. The Ni-hook contains the motif His-Cys-X-X-Pro-Cys-Gly-X-Tyr; it provides most of the interactions critical for metal binding and catalysis and is, therefore, a likely diagnostic of NiSODs.[11][12]
|
|
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
In higher plants, SOD isozymes have been localized in different cell compartments. Mn-SOD is present in mitochondria and peroxisomes. Fe-SOD has been found mainly in chloroplasts but has also been detected in peroxisomes, and CuZn-SOD has been localized in cytosol, chloroplasts, peroxisomes, and apoplast.[14][15]
Human
Three forms of superoxide dismutase are present in humans, in all other mammals, and most chordates. SOD1 is located in the cytoplasm, SOD2 in the mitochondria, and SOD3 is extracellular. The first is a dimer (consists of two units), whereas the others are tetramers (four subunits). SOD1 and SOD3 contain copper and zinc, whereas SOD2, the mitochondrial enzyme, has manganese in its reactive centre. The genes are located on chromosomes 21, 6, and 4, respectively (21q22.1, 6q25.3 and 4p15.3-p15.1).
|
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Plants
In higher plants, superoxide dismutase enzymes (SODs) act as antioxidants and protect cellular components from being oxidized by reactive oxygen species (ROS).[18] ROS can form as a result of drought, injury, herbicides and pesticides, ozone, plant metabolic activity, nutrient deficiencies, photoinhibition, temperature above and below ground, toxic metals, and UV or gamma rays.[19][20] To be specific, molecular O2 is reduced to O−
2 (a ROS called superoxide) when it absorbs an excited electron released from compounds of the electron transport chain. Superoxide is known to denature enzymes, oxidize lipids, and fragment DNA.[19] SODs catalyze the production of O2 and H
2O
2 from superoxide (O−
2), which results in less harmful reactants.
When acclimating to increased levels of oxidative stress, SOD concentrations typically increase with the degree of stress conditions. The compartmentalization of different forms of SOD throughout the plant makes them counteract stress very effectively. There are three well-known and -studied classes of SOD metallic coenzymes that exist in plants. First, Fe SODs consist of two species, one homodimer (containing 1–2 g Fe) and one tetramer (containing 2–4 g Fe). They are thought to be the most ancient SOD metalloenzymes and are found within both prokaryotes and eukaryotes. Fe SODs are most abundantly localized inside plant chloroplasts, where they are indigenous. Second, Mn SODs consist of a homodimer and homotetramer species each containing a single Mn(III) atom per subunit. They are found predominantly in mitochondrion and peroxisomes. Third, Cu-Zn SODs have electrical properties very different from those of the other two classes. These are concentrated in the chloroplast, cytosol, and in some cases the extracellular space. Note that Cu-Zn SODs provide less protection than Fe SODs when localized in the chloroplast.[18][19][20]
Bacteria
Human white blood cells use enzymes such as NADPH oxidase to generate superoxide and other reactive oxygen species to kill bacteria. During infection, some bacteria (e.g., Burkholderia pseudomallei) therefore produce superoxide dismutase to protect themselves from being killed.[21]
Biochemistry
SOD out-competes damaging reactions of superoxide, thus protecting the cell from superoxide toxicity.
The reaction of superoxide with non-radicals is spin-forbidden. In biological systems, this means that its main reactions are with itself (dismutation) or with another biological radical such as nitric oxide (NO) or with a transition-series metal. The superoxide anion radical (O−
2) spontaneously dismutes to O2 and hydrogen peroxide (H
2O
2) quite rapidly (~105 M−1s−1 at pH 7). SOD is necessary because superoxide reacts with sensitive and critical cellular targets. For example, it reacts with the NO radical, and makes toxic peroxynitrite.
Because the uncatalysed dismutation reaction for superoxide requires two superoxide molecules to react with each other, the dismutation rate is second-order with respect to initial superoxide concentration. Thus, the half-life of superoxide, although very short at high concentrations (e.g., 0.05 seconds at 0.1mM) is actually quite long at low concentrations (e.g., 14 hours at 0.1 nM). In contrast, the reaction of superoxide with SOD is first order with respect to superoxide concentration. Moreover, superoxide dismutase has the largest kcat/KM (an approximation of catalytic efficiency) of any known enzyme (~7 x 109 M−1s−1),[22] this reaction being limited only by the frequency of collision between itself and superoxide. That is, the reaction rate is "diffusion-limited".
The high efficiency of superoxide dismutase seems necessary: even at the subnanomolar concentrations achieved by the high concentrations of SOD within cells, superoxide inactivates the citric acid cycle enzyme aconitase, can poison energy metabolism, and releases potentially toxic iron. Aconitase is one of several iron-sulfur-containing (de)hydratases in metabolic pathways shown to be inactivated by superoxide.[23]
Stability and folding mechanism
SOD1 is an extremely stable protein. In the holo form (both copper and zinc bound) the melting point is > 90 °C. In the apo form (no copper or zinc bound) the melting point is ~60 °C.[24] By differential scanning calorimetry (DSC), holo SOD1 unfolds by a two-state mechanism: from dimer to two unfolded monomers.[24] In chemical denaturation experiments, holo SOD1 unfolds by a three-state mechanism with observation of a folded monomeric intermediate.[25]
Physiology
Superoxide is one of the main reactive oxygen species in the cell. As a consequence, SOD serves a key antioxidant role. The physiological importance of SODs is illustrated by the severe pathologies evident in mice genetically engineered to lack these enzymes. Mice lacking SOD2 die several days after birth, amid massive oxidative stress.[26] Mice lacking SOD1 develop a wide range of pathologies, including hepatocellular carcinoma,[27] an acceleration of age-related muscle mass loss,[28] an earlier incidence of cataracts, and a reduced lifespan. Mice lacking SOD3 do not show any obvious defects and exhibit a normal lifespan, though they are more sensitive to hyperoxic injury.[29] Knockout mice of any SOD enzyme are more sensitive to the lethal effects of superoxide-generating compounds, such as paraquat and diquat (herbicides).
Drosophila lacking SOD1 have a dramatically shortened lifespan, whereas flies lacking SOD2 die before birth. Depletion of SOD1 and SOD2 in the nervous system and muscles of Drosophila is associated with reduced lifespan.[30] The accumulation of neuronal and muscular ROS appears to contribute to age-associated impairments. When overexpression of mitochondrial SOD2 is induced, the lifespan of adult Drosophila is extended.[31]
Among black garden ants (Lasius niger), the lifespan of queens is an order of magnitude greater than of workers despite no systematic nucleotide sequence difference between them.[32] The SOD3 gene was found to be the most differentially over-expressed in the brains of queen vs worker ants. This finding raises the possibility of an important role of antioxidant function in modulating lifespan.[32]
SOD knockdowns in the worm C. elegans do not cause major physiological disruptions. However, the lifespan of C. elegans can be extended by superoxide/catalase mimetics suggesting that oxidative stress is a major determinant of the rate of aging.[33]
Knockout or null mutations in SOD1 are highly detrimental to aerobic growth in the budding yeast Saccharomyces cerevisiae and result in a dramatic reduction in post-diauxic lifespan. In wild-type S. cerevisiae, DNA damage rates increased 3-fold with age, but more than 5-fold in mutants deleted for either the SOD1 or SOD2 genes.[34] Reactive oxygen species levels increase with age in these mutant strains and show a similar pattern to the pattern of DNA damage increase with age. Thus it appears that superoxide dismutase plays a substantial role in preserving genome integrity during aging in S. cerevisiae. SOD2 knockout or null mutations cause growth inhibition on respiratory carbon sources in addition to decreased post-diauxic lifespan.
In the fission yeast Schizosaccharomyces pombe, deficiency of mitochondrial superoxide dismutase SOD2 accelerates chronological aging.[35]
Several prokaryotic SOD null mutants have been generated, including E. coli. The loss of periplasmic CuZnSOD causes loss of virulence and might be an attractive target for new antibiotics.
Role in disease
Mutations in the first SOD enzyme (SOD1) can cause familial amyotrophic lateral sclerosis (ALS, a form of motor neuron disease).[36][37][38][39] The most common mutation in the U.S. is A4V, while the most intensely studied is G93A. Inactivation of SOD1 causes hepatocellular carcinoma.[27] Diminished SOD3 activity has been linked to lung diseases such as acute respiratory distress syndrome (ARDS) or chronic obstructive pulmonary disease (COPD).[40][41][42] Superoxide dismutase is not expressed in neural crest cells in the developing fetus. Hence, high levels of free radicals can cause damage to them and induce dysraphic anomalies (neural tube defects).
Mutations in SOD1 can cause familial ALS (several pieces of evidence also show that wild-type SOD1, under conditions of cellular stress, is implicated in a significant fraction of sporadic ALS cases, which represent 90% of ALS patients.),[43] by a mechanism that is presently not understood, but not due to loss of enzymatic activity or a decrease in the conformational stability of the SOD1 protein. Overexpression of SOD1 has been linked to the neural disorders seen in Down syndrome.[44] In patients with thalassemia, SOD will increase as a form of compensation mechanism. However, in the chronic stage, SOD does not seem to be sufficient and tends to decrease due to the destruction of proteins from the massive reaction of oxidant-antioxidant.[45]
In mice, the extracellular superoxide dismutase (SOD3, ecSOD) contributes to the development of hypertension.[46][47] Inactivation of SOD2 in mice causes perinatal lethality.[26]
Medical uses
Supplementary superoxide dimutase has been suggested as a treatment to prevent bronchopulmonary dysplasia in infants who are born preterm, however the effectiveness of his treatment is not clear.[48]
Research
SOD has been used in experimental treatment of chronic inflammation in inflammatory bowl conditions.[49][50] SOD may ameliorate cis-platinum-induced nephrotoxicity (rodent studies).[51] As "Orgotein" or "ontosein", a pharmacologically-active purified bovine liver SOD, it is also effective in the treatment of urinary tract inflammatory disease in man.[52] For a time, bovine liver SOD even had regulatory approval in several European countries for such use. This was cut short by concerns about prion disease.
An SOD-mimetic agent, TEMPOL, is currently in clinical trials for radioprotection and to prevent radiation-induced dermatitis.[53] TEMPOL and similar SOD-mimetic nitroxides exhibit a multiplicity of actions in diseases involving oxidative stress.[54]
Cosmetic uses
SOD may reduce free radical damage to skin—for example, to reduce fibrosis following radiation for breast cancer. Studies of this kind must be regarded as tentative, however, as there were not adequate controls in the study including a lack of randomization, double-blinding, or placebo.[55] Superoxide dismutase is known to reverse fibrosis, possibly through de-differentiation of myofibroblasts back to fibroblasts.[56]
Commercial sources
SOD is commercially obtained from marine phytoplankton, bovine liver, horseradish, cantaloupe, and certain bacteria. For therapeutic purpose, SOD is usually injected locally. There is no evidence that ingestion of unprotected SOD or SOD-rich foods can have any physiological effects, as all ingested SOD is broken down into amino acids before being absorbed. However, ingestion of SOD bound to wheat proteins could improve its therapeutic activity, at least in theory.[57]
See also
- Catalase
- Glutathione peroxidase
- Jiaogulan
- NADPH oxidase, an enzyme which produces superoxide
- Peroxidase
References
- PDB: 1VAR; Borgstahl GE, Parge HE, Hickey MJ, Johnson MJ, Boissinot M, Hallewell RA, et al. (April 1996). "Human mitochondrial manganese superoxide dismutase polymorphic variant Ile58Thr reduces activity by destabilizing the tetrameric interface". Biochemistry. 35 (14): 4287–4297. doi:10.1021/bi951892w. PMID 8605177. S2CID 7450190.
- Hayyan M, Hashim MA, AlNashef IM (March 2016). "Superoxide Ion: Generation and Chemical Implications". Chemical Reviews. 116 (5): 3029–3085. doi:10.1021/acs.chemrev.5b00407. PMID 26875845.
- McCord JM, Fridovich I (November 1969). "Superoxide dismutase. An enzymic function for erythrocuprein (hemocuprein)". The Journal of Biological Chemistry. 244 (22): 6049–6055. doi:10.1016/S0021-9258(18)63504-5. PMID 5389100.
- McCord JM, Fridovich I (1988). "Superoxide dismutase: the first twenty years (1968–1988)". Free Radical Biology & Medicine. 5 (5–6): 363–369. doi:10.1016/0891-5849(88)90109-8. PMID 2855736.
- Brewer GJ (September 1967). "Achromatic regions of tetrazolium stained starch gels: inherited electrophoretic variation". American Journal of Human Genetics. 19 (5): 674–680. PMC 1706241. PMID 4292999.
- PDB: 2SOD;Tainer JA, Getzoff ED, Beem KM, Richardson JS, Richardson DC (September 1982). "Determination and analysis of the 2 A-structure of copper, zinc superoxide dismutase". Journal of Molecular Biology. 160 (2): 181–217. doi:10.1016/0022-2836(82)90174-7. PMID 7175933.
- Quint P, Reutzel R, Mikulski R, McKenna R, Silverman DN (February 2006). "Crystal structure of nitrated human manganese superoxide dismutase: mechanism of inactivation". Free Radical Biology & Medicine. 40 (3): 453–458. doi:10.1016/j.freeradbiomed.2005.08.045. PMID 16443160.
- Richardson J, Thomas KA, Rubin BH, Richardson DC (April 1975). "Crystal structure of bovine Cu,Zn superoxide dismutase at 3 A resolution: chain tracing and metal ligands". Proceedings of the National Academy of Sciences of the United States of America. 72 (4): 1349–1353. doi:10.1073/pnas.72.4.1349. PMC 432531. PMID 1055410..
- Tainer JA, Getzoff ED, Richardson JS, Richardson DC (1983). "Structure and mechanism of copper, zinc superoxide dismutase". Nature. 306 (5940): 284–287. Bibcode:1983Natur.306..284T. doi:10.1038/306284a0. PMID 6316150. S2CID 4266810.
- PDB: 1N0J; Borgstahl GE, Parge HE, Hickey MJ, Beyer WF, Hallewell RA, Tainer JA (October 1992). "The structure of human mitochondrial manganese superoxide dismutase reveals a novel tetrameric interface of two 4-helix bundles". Cell. 71 (1): 107–118. doi:10.1016/0092-8674(92)90270-M. PMID 1394426. S2CID 41611695.
- Barondeau DP, Kassmann CJ, Bruns CK, Tainer JA, Getzoff ED (June 2004). "Nickel superoxide dismutase structure and mechanism". Biochemistry. 43 (25): 8038–8047. doi:10.1021/bi0496081. PMID 15209499. S2CID 10700340.
- PDB: 1Q0M; Wuerges J, Lee JW, Yim YI, Yim HS, Kang SO, Djinovic Carugo K (June 2004). "Crystal structure of nickel-containing superoxide dismutase reveals another type of active site". Proceedings of the National Academy of Sciences of the United States of America. 101 (23): 8569–8574. Bibcode:2004PNAS..101.8569W. doi:10.1073/pnas.0308514101. PMC 423235. PMID 15173586.
- PDB: 1SDY; Djinović K, Gatti G, Coda A, Antolini L, Pelosi G, Desideri A, et al. (December 1991). "Structure solution and molecular dynamics refinement of the yeast Cu,Zn enzyme superoxide dismutase". Acta Crystallographica Section B: Structural Science. 47 ( Pt 6) (6): 918–927. doi:10.1107/S0108768191004949. PMID 1772629.
- Corpas FJ, Barroso JB, del Río LA (April 2001). "Peroxisomes as a source of reactive oxygen species and nitric oxide signal molecules in plant cells". Trends in Plant Science. 6 (4): 145–150. doi:10.1016/S1360-1385(01)01898-2. PMID 11286918.
- Corpas FJ, Fernández-Ocaña A, Carreras A, Valderrama R, Luque F, Esteban FJ, et al. (July 2006). "The expression of different superoxide dismutase forms is cell-type dependent in olive (Olea europaea L.) leaves". Plant & Cell Physiology. 47 (7): 984–994. doi:10.1093/pcp/pcj071. PMID 16766574.
- PDB: 3CQQ; Cao X, Antonyuk SV, Seetharaman SV, Whitson LJ, Taylor AB, Holloway SP, et al. (June 2008). "Structures of the G85R variant of SOD1 in familial amyotrophic lateral sclerosis". The Journal of Biological Chemistry. 283 (23): 16169–16177. doi:10.1074/jbc.M801522200. PMC 2414278. PMID 18378676.
- PDB: 2JLP; Antonyuk SV, Strange RW, Marklund SL, Hasnain SS (May 2009). "The structure of human extracellular copper-zinc superoxide dismutase at 1.7 A resolution: insights into heparin and collagen binding". Journal of Molecular Biology. 388 (2): 310–326. doi:10.1016/j.jmb.2009.03.026. PMID 19289127.
- Alscher RG, Erturk N, Heath LS (May 2002). "Role of superoxide dismutases (SODs) in controlling oxidative stress in plants". Journal of Experimental Botany. 53 (372): 1331–1341. doi:10.1093/jexbot/53.372.1331. PMID 11997379.
- Smirnoff N (September 1993). "The role of active oxygen in the response of plants to water deficit and desiccation". The New Phytologist. 125 (1): 27–58. doi:10.1111/j.1469-8137.1993.tb03863.x. PMID 33874604.
- Raychaudhuri SS, Deng XW (2008). "The Role of Superoxide Dismutase in Combating Oxidative Stress in Higher Plants". The Botanical Review. 66 (1): 89–98. doi:10.1007/BF02857783. S2CID 7663001.
- Vanaporn M, Wand M, Michell SL, Sarkar-Tyson M, Ireland P, Goldman S, et al. (August 2011). "Superoxide dismutase C is required for intracellular survival and virulence of Burkholderia pseudomallei". Microbiology. 157 (Pt 8): 2392–2400. doi:10.1099/mic.0.050823-0. PMID 21659326.
- Heinrich PC, Löffler G, Petrifies PE (2006). Biochemie und Pathobiochemie (Springer-Lehrbuch) (German ed.). Berlin: Springer. p. 123. ISBN 978-3-540-32680-9.
- Gardner PR, Raineri I, Epstein LB, White CW (June 1995). "Superoxide radical and iron modulate aconitase activity in mammalian cells". The Journal of Biological Chemistry. 270 (22): 13399–13405. doi:10.1074/jbc.270.22.13399. PMID 7768942.
- Stathopulos PB, Rumfeldt JA, Karbassi F, Siddall CA, Lepock JR, Meiering EM (March 2006). "Calorimetric analysis of thermodynamic stability and aggregation for apo and holo amyotrophic lateral sclerosis-associated Gly-93 mutants of superoxide dismutase". The Journal of Biological Chemistry. 281 (10): 6184–6193. doi:10.1074/jbc.M509496200. PMID 16407238.
- Rumfeldt JA, Stathopulos PB, Chakrabarrty A, Lepock JR, Meiering EM (January 2006). "Mechanism and thermodynamics of guanidinium chloride-induced denaturation of ALS-associated mutant Cu,Zn superoxide dismutases". Journal of Molecular Biology. 355 (1): 106–123. doi:10.1016/j.jmb.2005.10.042. PMID 16307756.
- Li Y, Huang TT, Carlson EJ, Melov S, Ursell PC, Olson JL, et al. (December 1995). "Dilated cardiomyopathy and neonatal lethality in mutant mice lacking manganese superoxide dismutase". Nature Genetics. 11 (4): 376–381. doi:10.1038/ng1295-376. PMID 7493016. S2CID 10900822.
- Elchuri S, Oberley TD, Qi W, Eisenstein RS, Jackson Roberts L, Van Remmen H, et al. (January 2005). "CuZnSOD deficiency leads to persistent and widespread oxidative damage and hepatocarcinogenesis later in life". Oncogene. 24 (3): 367–380. doi:10.1038/sj.onc.1208207. PMID 15531919.
- Muller FL, Song W, Liu Y, Chaudhuri A, Pieke-Dahl S, Strong R, et al. (June 2006). "Absence of CuZn superoxide dismutase leads to elevated oxidative stress and acceleration of age-dependent skeletal muscle atrophy". Free Radical Biology & Medicine. 40 (11): 1993–2004. doi:10.1016/j.freeradbiomed.2006.01.036. PMID 16716900.
- Sentman ML, Granström M, Jakobson H, Reaume A, Basu S, Marklund SL (March 2006). "Phenotypes of mice lacking extracellular superoxide dismutase and copper- and zinc-containing superoxide dismutase". The Journal of Biological Chemistry. 281 (11): 6904–6909. doi:10.1074/jbc.M510764200. PMID 16377630.
- Oka S, Hirai J, Yasukawa T, Nakahara Y, Inoue YH (August 2015). "A correlation of reactive oxygen species accumulation by depletion of superoxide dismutases with age-dependent impairment in the nervous system and muscles of Drosophila adults". Biogerontology. 16 (4): 485–501. doi:10.1007/s10522-015-9570-3. PMID 25801590. S2CID 18050827.
- Sun J, Folk D, Bradley TJ, Tower J (June 2002). "Induced overexpression of mitochondrial Mn-superoxide dismutase extends the life span of adult Drosophila melanogaster". Genetics. 161 (2): 661–672. doi:10.1093/genetics/161.2.661. PMC 1462135. PMID 12072463.
- Lucas ER, Keller L (July 2018). "Elevated expression of ageing and immunity genes in queens of the black garden ant". Experimental Gerontology. 108: 92–98. doi:10.1016/j.exger.2018.03.020. PMID 29625209. S2CID 5045743.
- Melov S, Ravenscroft J, Malik S, Gill MS, Walker DW, Clayton PE, et al. (September 2000). "Extension of life-span with superoxide dismutase/catalase mimetics". Science. 289 (5484): 1567–1569. Bibcode:2000Sci...289.1567M. doi:10.1126/science.289.5484.1567. PMID 10968795. S2CID 21519801.
- Muid KA, Karakaya HÇ, Koc A (February 2014). "Absence of superoxide dismutase activity causes nuclear DNA fragmentation during the aging process". Biochemical and Biophysical Research Communications. 444 (2): 260–263. doi:10.1016/j.bbrc.2014.01.056. hdl:11147/5542. PMID 24462872.
- Ogata T, Senoo T, Kawano S, Ikeda S (January 2016). "Mitochondrial superoxide dismutase deficiency accelerates chronological aging in the fission yeast Schizosaccharomyces pombe". Cell Biology International. 40 (1): 100–106. doi:10.1002/cbin.10556. PMID 26507459. S2CID 205563521.
- Milani P, Gagliardi S, Cova E, Cereda C (2011). "SOD1 Transcriptional and Posttranscriptional Regulation and Its Potential Implications in ALS". Neurology Research International. 2011: 458427. doi:10.1155/2011/458427. PMC 3096450. PMID 21603028.
- Deng HX, Hentati A, Tainer JA, Iqbal Z, Cayabyab A, Hung WY, et al. (August 1993). "Amyotrophic lateral sclerosis and structural defects in Cu,Zn superoxide dismutase". Science. 261 (5124): 1047–1051. Bibcode:1993Sci...261.1047D. doi:10.1126/science.8351519. PMID 8351519.
- Conwit RA (December 2006). "Preventing familial ALS: a clinical trial may be feasible but is an efficacy trial warranted?". Journal of the Neurological Sciences. 251 (1–2): 1–2. doi:10.1016/j.jns.2006.07.009. PMID 17070848. S2CID 33105812.
- Al-Chalabi A, Leigh PN (August 2000). "Recent advances in amyotrophic lateral sclerosis". Current Opinion in Neurology. 13 (4): 397–405. doi:10.1097/00019052-200008000-00006. PMID 10970056. S2CID 21577500.
- Young RP, Hopkins R, Black PN, Eddy C, Wu L, Gamble GD, et al. (May 2006). "Functional variants of antioxidant genes in smokers with COPD and in those with normal lung function". Thorax. 61 (5): 394–399. doi:10.1136/thx.2005.048512. PMC 2111196. PMID 16467073.
- Ganguly K, Depner M, Fattman C, Bein K, Oury TD, Wesselkamper SC, et al. (May 2009). "Superoxide dismutase 3, extracellular (SOD3) variants and lung function". Physiological Genomics. 37 (3): 260–267. doi:10.1152/physiolgenomics.90363.2008. PMC 2685504. PMID 19318538.
- Gongora MC, Lob HE, Landmesser U, Guzik TJ, Martin WD, Ozumi K, et al. (October 2008). "Loss of extracellular superoxide dismutase leads to acute lung damage in the presence of ambient air: a potential mechanism underlying adult respiratory distress syndrome". The American Journal of Pathology. 173 (4): 915–926. doi:10.2353/ajpath.2008.080119. PMC 2543061. PMID 18787098.
- Gagliardi S, Cova E, Davin A, Guareschi S, Abel K, Alvisi E, et al. (August 2010). "SOD1 mRNA expression in sporadic amyotrophic lateral sclerosis". Neurobiology of Disease. 39 (2): 198–203. doi:10.1016/j.nbd.2010.04.008. PMID 20399857. S2CID 207065284.
- Groner Y, Elroy-Stein O, Avraham KB, Schickler M, Knobler H, Minc-Golomb D, et al. (1994). "Cell damage by excess CuZnSOD and Down's syndrome". Biomedicine & Pharmacotherapy. 48 (5–6): 231–240. doi:10.1016/0753-3322(94)90138-4. PMID 7999984.
- Rujito L, Mulatsih S, Sofro AS (May 2015). "Status of Superoxide Dismutase in Transfusion Dependent Thalassaemia". North American Journal of Medical Sciences. 7 (5): 194–198. doi:10.4103/1947-2714.157480. PMC 4462814. PMID 26110130.
- Gongora MC, Qin Z, Laude K, Kim HW, McCann L, Folz JR, et al. (September 2006). "Role of extracellular superoxide dismutase in hypertension". Hypertension. 48 (3): 473–481. doi:10.1161/01.HYP.0000235682.47673.ab. PMID 16864745.
- Lob HE, Marvar PJ, Guzik TJ, Sharma S, McCann LA, Weyand C, et al. (February 2010). "Induction of hypertension and peripheral inflammation by reduction of extracellular superoxide dismutase in the central nervous system". Hypertension. 55 (2): 277–83, 6p following 283. doi:10.1161/HYPERTENSIONAHA.109.142646. PMC 2813894. PMID 20008675.
- Albertella M, Gentyala RR, Paraskevas T, Ehret D, Bruschettini M, Soll R, et al. (Cochrane Neonatal Group) (October 2023). "Superoxide dismutase for bronchopulmonary dysplasia in preterm infants". The Cochrane Database of Systematic Reviews. 10 (10): CD013232. doi:10.1002/14651858.CD013232.pub2. PMC 10561150. PMID 37811631.
- Seguí J, Gironella M, Sans M, Granell S, Gil F, Gimeno M, et al. (September 2004). "Superoxide dismutase ameliorates TNBS-induced colitis by reducing oxidative stress, adhesion molecule expression, and leukocyte recruitment into the inflamed intestine". Journal of Leukocyte Biology. 76 (3): 537–544. doi:10.1189/jlb.0304196. PMID 15197232. S2CID 15028921.
- Seguí J, Gironella M, Sans M, Granell S, Gil F, Gimeno M, et al. (September 2004). "Superoxide dismutase ameliorates TNBS-induced colitis by reducing oxidative stress, adhesion molecule expression, and leukocyte recruitment into the inflamed intestine". Journal of Leukocyte Biology. 76 (3): 537–544. doi:10.1189/jlb.0304196. PMID 15197232. S2CID 15028921.
- McGinness JE, Proctor PH, Demopoulos HB, Hokanson JA, Kirkpatrick DS (1978). "Amelioration of cis-platinum nephrotoxicity by orgotein (superoxide dismutase)". Physiological Chemistry and Physics. 10 (3): 267–277. PMID 733940.
- Marberger H, Huber W, Bartsch G, Schulte T, Swoboda P (1974). "Orgotein: a new antiinflammatory metalloprotein drug evaluation of clinical efficacy and safety in inflammatory conditions of the urinary tract". International Urology and Nephrology. 6 (2): 61–74. doi:10.1007/bf02081999. PMID 4615073. S2CID 23880216.
- Clinical trial number NCT01324141 for "Topical MTS-01 for Dermatitis During Radiation and Chemotherapy for Anal Cancer" at ClinicalTrials.gov
- Wilcox CS (May 2010). "Effects of tempol and redox-cycling nitroxides in models of oxidative stress". Pharmacology & Therapeutics. 126 (2): 119–145. doi:10.1016/j.pharmthera.2010.01.003. PMC 2854323. PMID 20153367.
- Campana F, Zervoudis S, Perdereau B, Gez E, Fourquet A, Badiu C, et al. (2004). "Topical superoxide dismutase reduces post-irradiation breast cancer fibrosis". Journal of Cellular and Molecular Medicine. 8 (1): 109–116. CiteSeerX 10.1.1.336.8033. doi:10.1111/j.1582-4934.2004.tb00265.x. PMC 6740277. PMID 15090266.
- Vozenin-Brotons MC, Sivan V, Gault N, Renard C, Geffrotin C, Delanian S, et al. (January 2001). "Antifibrotic action of Cu/Zn SOD is mediated by TGF-beta1 repression and phenotypic reversion of myofibroblasts". Free Radical Biology & Medicine. 30 (1): 30–42. doi:10.1016/S0891-5849(00)00431-7. PMID 11134893.
- Romao S (March 2015). "Therapeutic value of oral supplementation with melon superoxide dismutase and wheat gliadin combination". Nutrition. 31 (3): 430–436. doi:10.1016/j.nut.2014.10.006. PMID 25701330.
External links
- Online Mendelian Inheritance in Man (OMIM): 105400 (ALS)
- The ALS Online Database
- A short but substantive overview of SOD and its literature.
- Damage-Based Theories of Aging Includes a discussion of the roles of SOD1 and SOD2 in aging.
- Physicians' Comm. For Responsible Med.
- SOD and Oxidative Stress Pathway Image
- PDBe-KB provides an overview of all the structure information available in the PDB for Human Superoxide dismutase [Cu-Zn]
- PDBe-KB provides an overview of all the structure information available in the PDB for Human Superoxide dismutase [Mn], mitochondrial
- PDBe-KB provides an overview of all the structure information available in the PDB for Human Extracellular superoxide dismutase [Cu-Zn]
| pyrazolones / pyrazolidines | |
|---|---|
| salicylates | |
| acetic acid derivatives and related substances | |
| oxicams |
|
| propionic acid derivatives (profens) |
|
| n-arylanthranilic acids (fenamates) | |
| COX-2 inhibitors (coxibs) | |
| other | |
| NSAID combinations | |
Key: underline indicates initially developed first-in-class compound of specific group; #WHO-Essential Medicines; †withdrawn drugs; ‡veterinary use. | |