Dilophosaurus wetherilli
Dilophosaurus wetherilli (gr. "lagarto de dos crestas de John Wetherill") es la única especie conocida del género extinto Dilophosaurus de dinosaurio terópodo dilofosáurido que vivió a principios del periodo Jurásico, hace aproximadamente 200 y 190 millones de años, entre el Sinemuriense y Pliensbachiense, en lo que hoy es Norteamérica. La especie Dilophosaurus sinensis, que anteriormente era considerada como perteneciente a este género, ha sido reclasificada como perteneciente al sinónimo más moderno del género Sinosaurus, como S. sinensis.[1]
| Dilophosaurus wetherilli | ||
|---|---|---|
| Rango temporal: 193 Ma Jurásico Inferior (Sinemuriense) | ||
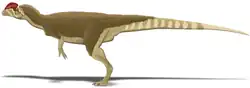 Recreación | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Theropoda | |
| (sin rango): | Neotheropoda | |
| Superfamilia: | Coelophysoidea | |
| Familia: | Dilophosauridae | |
| Género: |
Dilophosaurus Welles, 1970 | |
| Especie: |
D. wetherilli (Welles, 1954) | |
| Sinonimia | ||
| ||
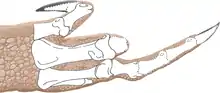
Con aproximadamente 7 metros de longitud, con un peso de aproximadamente 400 kilogramos, Dilophosaurus fue uno de los primeros dinosaurios depredadores grandes, aunque más pequeño que algunos terópodos posteriores. Era delgado y de constitución ligera, y el cráneo era proporcionalmente grande, pero delicado. El hocico era estrecho y la mandíbula superior tenía una brecha de la ventana de la nariz. Tenía un par de crestas longitudinales en forma de placa en su cráneo, similar a un casuario con dos crestas. La mandíbula era delgada y delicada en la parte delantera, pero profunda en la parte posterior. Los dientes eran largos, curvados, delgados y comprimidos hacia los lados. Los de la mandíbula inferior eran mucho más pequeños que los de la mandíbula superior. La mayoría de los dientes tenían dientes en sus bordes frontal y posterior. El cuello era largo, y sus vértebras eran huecas, y muy ligeras. Los brazos eran poderosos, con un hueso del brazo largo y delgado. Las manos tenían cuatro dedos: el primero era corto pero fuerte y tenía una garra grande, los dos dedos siguientes eran más largos y delgados con garras más pequeñas, y el cuarto era vestigial. El hueso del muslo era enorme, los pies fuertes y los dedos del pie con grandes garras.
Los paleontólogos, consideraron que el Dilophosaurus podría ser un género basal que generó clados como los ceratosaurios y los tetanuros. Otros paleontólogos, piensan al contrario que en realidad este género no es más que un celofísido de gran tamaño. Los primeros especímenes fueron descritos en 1954, pero no fue hasta una década más tarde que el género recibió su nombre actual. Dilophosaurus es uno de los terópodos jurásicos más antiguos conocidos, pero también uno de los menos comprendidos. Hoy es considerado un miembro de la familia Dilophosauridae junto con Dracovenator, un grupo ubicado entre los celofísidos y los terópodos posteriores. Dilophosaurus habría sido activo y bípedo, y puede haber cazado animales grandes y también podría haberse alimentado de animales y peces más pequeños. La función de las crestas es desconocida, eran demasiado débiles para la batalla, pero pueden haber sido utilizados en la visualización, como el reconocimiento de especies y la selección sexual. Puede haber crecido rápidamente, alcanzando una tasa de crecimiento de 30 a 35 kilogramos por año al comienzo de su vida. El espécimen holotipo tenía múltiples paleopatologías, incluyendo lesiones cicatrizadas y signos de una anomalía del desarrollo. El Dilophosaurus es conocido de la Formación Kayenta, y vivió junto a dinosaurios como Megapnosaurus y Sarahsaurus.
El Dilophosaurus ha sido representado en diferentes ocasiones en la cultura popular. Su aparición más famosa se produjo en la película Parque Jurásico, de 1993. Aunque con brillantes efectos especiales, esta película representa a Dilophosaurus muchísimo más pequeño de lo que era en realidad, además de que la película le atribuye la capacidad de desplegar un collar extensible, como el lagarto agámido Chlamydosaurus kingii y también la capacidad de proyectar veneno sobre sus presas. La capacidad de proyectar veneno ya estaba presente en la descripción que de esta especie hacía la novela en la que se basa la película. Estas dos últimas características provienen de la imaginación de los autores, dado que no hay forma de comprobar que este dinosaurio haya poseído un collar extensible (solo para el film) o la capacidad de escupir veneno. Fue designado como el dinosaurio estatal de Connecticut en 2017.
Descripción
El Dilophosaurus fue uno de los primeros dinosaurios depredadores grandes, un terópodo de tamaño mediano, aunque pequeño en comparación con algunos de los terópodos posteriores.[2][3] Esbelto y de constitución ligera, su tamaño era comparable al de un oso pardo.[3][4][5] El espécimen más grande conocido pesaba unos 400 kilogramos, medía unos 7 metros de largo, y su cráneo tenía 590 milímetros de largo. El espécimen holotipo es más pequeño, pesaba aproximadamente 283 kilogramos, tenía 6.03 metros de largo, con una altura de cadera de aproximadamente 1,36 metros y su cráneo tenía 523 milímetros de largo.[4][6] Algunos investigadores han interpretado que una huella de descanso de un terópodo similar al Dilophosaurus y Liliensternus muestra impresiones de plumas alrededor del vientre y los pies, similar al plumón.[7][8] Otros investigadores interpretan estas impresiones como artefactos sedimentológicos creados a medida que el dinosaurio se movía, aunque esta interpretación no descarta que el creador de huellas haya tenido plumas.[9][10]
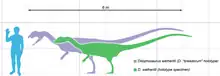
La característica más distintiva del Dilophosaurus es el par de crestas redondeadas en su cráneo, usado posiblemente para la exhibición.[2][11] En 1997, Dodson notó que las crestas craneales aparecieron antes en el Dilophosaurus que en otros tipos de terópodos.[12] Estudios de Robert Gay demuestran que pudo haber habido variación de tamaño entre ambos sexos, pero nada dicen acerca de la cresta en sí.[13] Los dientes del Dilophosaurus son largos, pero tienen una base bastante pequeña y se amplían basalmente.[14] Otra característica del cráneo era una muesca detrás de la primera fila de dientes, dando al Dilophosaurus casi un aspecto de cocodrilo, similar a los dinosaurios supuestamente piscívoros espinosáuridos. Esta "diastema" existió debido una conexión débil entre los huesos premaxilares y maxilares del cráneo. Esta conformación llevó a la hipótesis temprana que el Dilophosaurus era un carroñero, con los dientes delanteros siendo demasiado débiles para derribar y sostener una presa grande.[15]
Esqueleto axial
El Dilophosaurus tenía 10 vértebras cervicales en el cuello, 14 dorsales en la espalda y 45 caudales en la cola. Tenía un cuello largo, que probablemente estaba flexionado casi 90° por el cráneo y por el hombro, sosteniendo el cráneo en una postura horizontal. Las vértebras cervicales eran inusualmente ligeras; sus centros, los "cuerpos" de las vértebras, estaban ahuecados por pleurocoelos, depresiones en los lados y centroscoelos, cavidades en el interior. Los arcos de las vértebras cervicales también tenían chonosis, huecos cónicos tan grandes que los huesos que los separaban eran a veces finos como el papel. El centro era plano-cóncavo, plano a débilmente convexo en la parte frontal y profundamente ahuecado o cóncavo en la parte posterior, similar a Ceratosaurus. Esto indica que el cuello era flexible, aunque tenía costillas cervicales superpuestas que estaban fusionadas con el centro. Las costillas cervicales eran delgadas y pueden haberse doblado fácilmente. El hueso atlas, la primera vértebra cervical que se une al cráneo, tenía un centro pequeño y cúbico y tenía una concavidad en el frente donde formaba una copa para el cóndilo occipital, protuberancia que conecta con la vértebra del atlas, en la parte posterior de la calavera. El hueso axis, la segunda vértebra cervical, tenía una espina pesada, y sus postzigópofisis, los procesos de las vértebras que se articulaban con las vértebras siguientes, se encontraron con prezygapofisis largas que se curvaban hacia arriba desde la tercera vértebra cervical. Los centros y espinas de las vértebras cervicales eran largas y bajas, y las espinas tenían tapas que daban la apariencia de una cruz de Malta, cruciforme, cuando se ve desde arriba, una característica distintiva de este dinosaurio. Las espinas neurales de las vértebras dorsal también eran bajas y se expandieron hacia adelante y hacia atrás, lo que formó grandes sitios de inserción para los ligamentos. Las vértebras sacras que ocuparon la longitud del ilion que no parecía estar fusionado. El centro de las vértebras caudales era muy consistente en longitud, pero su diámetro se hacía más pequeño hacia la parte posterior, y pasaron de elíptico a circular en sección transversal.[2][16]
Miembros
Las escápulas eran de longitud moderada y cóncavas en sus lados internos para seguir la curvatura del cuerpo. Los omóplatos eran anchos, particularmente la parte superior, que era rectangular o cuadrada, una característica única. Los coracoides eran elípticos y no se fusionaron con las escápulas. Los brazos eran poderosos y tenían hoyos profundos y procesos fuertes para la unión de músculos y ligamentos. El húmero, el hueso del brazo, era grande y delgado, con epipodiales fuertes y el cúbito, hueso del antebrazo, era grueso y recto, con un olécranon fuerte. Las manos tenían cuatro dedos, el primero era más corto pero más fuerte que los dos dedos siguientes, con una garra grande y los dos dedos siguientes eran más largos y delgados, con garras más pequeñas. El tercer dedo se redujo, y el cuarto fue vestigial, retenido, pero sin función. La cresta del ilion era más alta sobre el pedúnculo ilíaco, el proceso hacia abajo del ilion y su lado externo era cóncavo. El pie del hueso púbico estaba ligeramente expandido, mientras que el extremo inferior estaba mucho más expandido en el isquion, que también tenía un eje muy delgado. Las patas traseras eran grandes, con un fémur, hueso del muslo, más largo que la tibia, hueso de la parte inferior de la pierna, lo contrario de, por ejemplo, Coelophysis. El fémur fue masivo, su eje era de forma sigmoidea, curvado como una S y su trocánter mayor estaba centrado en el eje. La tibia tenía una tuberosidad desarrollada y se expandió en el extremo inferior. El hueso del astrágalo, hueso del tobillo, se separó de la tibia y el calcáneo, y formó la mitad del alveolo para el peroné. Tenía pies largos y robustos con tres dedos bien desarrollados que tenían grandes garras. El tercer dedo fue el más fuerte, y el primer dedo del pie más pequeño, el hallux, se mantuvo sin tocar el suelo.[2][16][17]
Cráneo
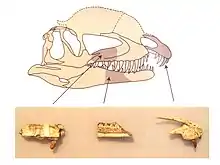
El cráneo del Dilophosaurus era grande en proporción al esqueleto general, pero delicado. El hocico era estrecho visto de frontal, cada vez más estrecho hacia la parte superior redondeada. El premaxilar, hueso frontal de la mandíbula superior, era largo y bajo cuando se ve desde un lado, y su superficie externa se volvió menos convexa desde el hocico hasta la narina, fosa nasal. Las fosas nasales se colocaron más atrás que en la mayoría de los otros terópodos. El premaxilar estaba débilmente unida al maxilar, el siguiente hueso de la mandíbula superior, solo se conectaba a la mitad del paladar, sin conexión en el costado. Hacia abajo, la premaxilar formó una pared para un espacio entre ella y el maxilar llamado espacio subnarial, también denominado "pliegue". Tal brecha también está presente en celofisoides. La brecha subnarial dio como resultado una diastema, un espacio en la fila de dientes, que también se ha denominado "muesca". Dentro de la brecha subnarial había una excavación profunda detrás de los dientes premaxilares, llamada fosa subnarial, que estaba amurallada por una quilla hacia abajo del premaxilar. La superficie externa del premaxilar estaba cubierta de foramenes de diferentes tamaños. La parte superior de los dos procesos premaxilares que se extendían hacia atrás era larga y baja y formaba la mayor parte del borde superior de las narinas alargadas. Se metía hacia adentro, lo que hizo que el área fuera cóncava en su perfil. La parte inferior del premaxilar que contiene los alveolos dentarios era ovalada. El maxilar era poco profundo, y estaba deprimido alrededor de la fenestra antorbital, una abertura grande en frente del ojo, formando un hueco redondeado hacia el frente y más liso que el resto del maxilar superior. Un foramen llamado fenestra preantorbital se abrió en este hueco en la curva delantera. Grandes forámenes corrían en el lado del maxilar, arriba de los alvéolos. Un profundo surco de nutrientes corría hacia atrás desde la foso subnarial a lo largo de la base de las placas interdentales del maxilar superior.[2][4][16][18]

El Dilophosaurus tenía un par de crestas altas, delgadas y arqueadas o en forma de placa longitudinalmente en el techo del cráneo. Las crestas se formaron significativamente por los huesos lagrimales y parcialmente por los huesos nasales. Las crestas se extendían hacia el techo del cráneo y daban la apariencia de un casuario con dos crestas. Como característica distintiva, cada cresta también tenía una proyección hacia atrás similar a un dedo. La superficie superior del hueso nasal entre las crestas era cóncava, y la parte nasal de la cresta se superponía a la parte lagrimal. Como solo un espécimen preserva la forma de las crestas, se desconoce si difieren en otros individuos. El hueso lagrimal tenía un borde superior engrosado único, donde formaba el borde superior en la parte posterior de la fenestra anterorbital. El hueso prefrontal formó el techo de la órbita ocular y tenía una barra en forma de L que hacía que parte de la superficie superior de la órbita fuera cóncava. La órbita era ovalada, y estrecha hacia la parte inferior. El hueso yugal tenía dos procesos apuntando hacia arriba, el primero de los cuales formaba parte del margen inferior de la fenestra anterorbital y parte del margen inferior de la órbita. Una proyección del hueso cuadrado en la fenestra temporal lateral, la abertura detrás del ojo, le dio a este un contorno reniforme, en forma de riñón. El foramen magnum, la gran abertura en la parte posterior de la caja craneana, tenía aproximadamente la mitad del ancho del cóndilo occipital, que era cordiforme, en forma de corazón, y tenía un cuello corto y una ranura en el costado.[2][3][18][19][20]
Mandíbula y dientes
La mandíbula era delgada y delicada en el frente, pero la región articular, donde se conectaba con el cráneo, era fuerte y la mandíbula era profunda alrededor de la fenestra mandibular, una abertura en su costado. La fenestra mandibular era pequeña en el Dilophosaurus, en comparación con la de los celofisoides. El proceso retroarticular de la mandíbula, una proyección hacia atrás, era largo, y la plataforma surangular era fuertemente horizontal. El hueso dentario, la parte frontal de la mandíbula, donde se unieron la mayoría de los dientes, tenía una barbilla curvada en lugar de puntiaguda. El mentón tenía un foramen grande en la punta, y una hilera de pequeños agujeros corrían en paralelo aproximado con el borde superior del dentario. En el lado interno, la sínfisis mandibular, donde se conectaban las dos mitades de la mandíbula inferior, era plana y lisa, y no mostraba signos de estar fusionada con su mitad opuesta. Un foramen de Meckelian corría a lo largo del lado externo del dentario.[2][16]

El Dilophosaurus tenía cuatro dientes en cada premaxilar, doce en cada maxilar y diecisiete en cada dentario. Los dientes generalmente eran largos, delgados y recurvados, con bases relativamente pequeñas. Estaban comprimidos de lado, ovales en sección transversal en la base, lenticulares, en forma de lente, arriba, y ligeramente cóncavos en sus lados externo e interno. El diente más grande del maxilar estaba o en el cuarto alveolo o cerca de él, y la altura de las coronas dentales disminuyó en las zonas traseras. El primer diente del maxilar apuntaba ligeramente hacia adelante desde su alveolo porque el borde inferior del proceso prexamilar, que se proyectaba hacia el maxilar, se volteaba hacia arriba. Los dientes del dentario eran mucho más pequeños que los del maxilar. El tercer o cuarto diente en el dentario del Dilophosaurus y algunos celofisoides eran los más grandes y parece haber encajado en la brecha subnarial de la mandíbula superior. La mayoría de los dientes tenían estrías en los bordes anterior y posterior, que se compensaban con surcos verticales, y eran más pequeños en la parte delantera. Hubo 31 a 41 dentículos en los bordes delanteros, y 29 a 33 en la parte posterior. Al menos el segundo y el tercer diente del premaxilar tenían dentículos, pero el cuarto diente no. Los dientes fueron cubiertos en una capa delgada de esmalte, 0.1 a 0.15 milímetros de espesor, que se extendía más allá de sus bases. Los alvéolos eran elípticos a casi circulares, y todos eran más grandes que las bases de los dientes que contenían, por lo que, se podían haber sostenido libremente en las mandíbulas. Aunque el número de alvéolos en el dentario parece indicar que los dientes estaban muy apretados, estaban bastante separados, debido al mayor tamaño de sus alvéolos. Las mandíbulas contenían dientes de reemplazo en varias etapas de la erupción. Las placas interdentales entre los dientes eran muy bajas.[2][16][14]
Descubrimiento e investigación
Los primeros especímenes de Dilophosaurus wetherilli fueron descritos en 1954, pero no fue hasta una década más tarde que el género recibió su nombre actual. Es conocido de la formación Kayenta, y vivió junto a dinosaurios como Megapnosaurus y Sarahsaurus ente otros durante el jurásico temprano, hace aproximadamente 193 millones de años.
Primeros hallazgos
En el verano de 1942, el paleontólogo estadounidense Charles L. Camp dirigió una expedición de campo del Museo de Paleontología de la Universidad de California (UCMP) en busca de vertebrados fósiles en el condado de Navajo en el norte de Arizona. La noticia de esto se extendió entre los nativos americanos navajos de allí, y Jesse Williams llevó a tres miembros de la expedición a algunos huesos fósiles que había descubierto en 1940. El área era parte de la formación Kayenta, a unos 32 kilómetros al norte de Cameron cerca de la ciudad de Tuba en la reserva india Navajo. Se encontraron tres esqueletos de dinosaurios en una lutita violácea, dispuestos en un triángulo, de unos 9.1 metros de largo por lado. El primero estaba casi completo, careciendo solo del frente del cráneo, partes de la pelvis y algunas vértebras. El segundo estaba muy erosionado, incluido el frente del cráneo, las mandíbulas inferiores, algunas vértebras, huesos de las extremidades y una mano articulada. El tercero estaba tan erosionado que solo consistía en fragmentos de vértebras. El primer esqueleto bueno estaba encerrado en un bloque de yeso después de diez días de trabajo y cargado en un camión, el segundo esqueleto se recogió fácilmente ya que casi se capeó por completo del suelo, pero el tercer esqueleto casi se había perdido.[2][11][21]

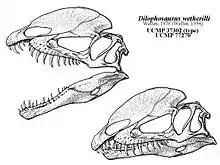
El primer espécimen casi completo fue limpiado y montado en la UCMP bajo la supervisión del paleontólogo estadounidense Wann Langston Jr., un proceso que le llevó a tres hombres dos años de trabajo. El esqueleto estaba montado en la pared en bajorrelieve, con la cola curvada hacia arriba, el cuello enderezado y la pierna izquierda movida hacia arriba para dar visibilidad, pero el resto del esqueleto se mantuvo en su posición de enterramiento. Como el cráneo fue aplastado, fue reconstruido con base en la parte posterior del cráneo del primer espécimen y al frente del segundo. La pelvis fue reconstruida como la de Allosaurus, y los pies también fueron reconstruidos. En ese momento, era uno de los esqueletos mejor conservados de un dinosaurio terópodo, aunque incompleto. En 1954, el paleontólogo estadounidense Samuel P. Welles, quien fue parte del grupo que excavó los esqueletos, describió preliminarmente y llamó a este dinosaurio como una nueva especie en el género existente Megalosaurus, M. wetherilli. El espécimen casi completo, catalogado como UCMP 37302, se convirtió en el holotipo, y el segundo espécimen,UCMP 37303 se incluyó en el hipodígmen, la muestra de especímenes que define un taxón, de la especie. El nombre específico honró a John Wetherill, un concejal navajo a quien Welles describió como un "explorador, amigo de los científicos y comerciante de confianza". Fue el sobrino de Wetherill, Milton, quien primero informó a la expedición de los fósiles. Welles colocó la nueva especie en Megalosaurus debido a las proporciones similares de los miembros con M. bucklandii, y porque no encontró grandes diferencias entre ellos. En ese momento, se usó Megalosaurus como un "taxón cajón de sastre", en el que se colocaron muchas especies de terópodos, independientemente de su edad o localidad.[2][3][11][22][23]
Espécimen de 1964
Welles regresó a la ciudad de Tuba en 1964 para determinar la edad de la formación Kayenta, se había sugerido que era del Triásico Tardío en edad, mientras que Welles pensó que era Jurásico Temprano o Medio, y descubrió otro esqueleto a 402,3 kilómetros al sur de donde se encontraron los ejemplares de 1942. El espécimen casi completo, catalogado como UCMP 77270 fue recolectado con la ayuda de William Breed del Museo del Norte de Arizona y otros. Durante la preparación de este espécimen, quedó claro que era un individuo más grande de M. wetherilli y que habría tenido dos crestas en la parte superior de su cráneo. Siendo un delgado plato de hueso, originalmente se creía que una cresta formaba parte del lado izquierdo faltante del cráneo que un carroñero había sacado de su posición. Cuando se hizo evidente que se trataba de una cresta, también se descubrió que habría una cresta correspondiente en el lado izquierdo, ya que la cresta derecha estaba a la derecha de la línea media, y era cóncava a lo largo de su longitud media. Este descubrimiento condujo a la reexaminación del espécimen de holotipo, que se encontró que tenía bases de dos huesos delgados, extendidos hacia arriba, que se rompieron juntas. Estas también representaban las crestas, pero anteriormente se había supuesto que formaban parte de un pómulo mal colocado. También se concluyó que los dos especímenes de 1942 eran juveniles, mientras que el espécimen de 1964 era un adulto, aproximadamente un tercio más grande que los demás.[2][18][22][14]
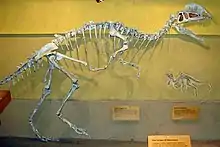
Posteriormente, Welles y un asistente corrigieron la montura de la pared del holotipo en función del nuevo esqueleto, restaurando las crestas, rehaciendo la pelvis, alargando las costillas del cuello y acercándolas más. Después de estudiar los esqueletos de los terópodos norteamericanos y europeos, Welles se dio cuenta de que el dinosaurio no pertenecía a Megalosaurus y necesitaba un nuevo nombre de género. En este momento, no se conocían otros terópodos con grandes crestas longitudales en sus cabezas, y el dinosaurio había ganado el interés de los paleontólogos. Se hizo un molde del holotipo y se distribuyeron moldes de fibra de vidrio en varias exhibiciones. Para facilitar el etiquetado de estos moldes, Welles decidió nombrar el nuevo género en una nota breve, en lugar de esperar hasta la publicación de una descripción detallada. En 1970 Welles acuñó el nuevo nombre de género Dilophosaurus, de las palabras griegas δι', di, que significa "dos", λόφος, lophos que significa "cresta" y σαυρος sauros que significa "lagarto" , "lagarto de dos crestas". Welles publicó una descripción osteológica detallada de Dilophosaurus en 1984, pero el espécimen de 1964 todavía no se ha descrito adecuadamente.[2][18][20][22][24] El Dilophosaurus fue el primer terópodo conocido del Jurásico temprano y sigue siendo uno de los ejemplos mejor conservados de esa edad.[3]
Nuevos hallazgos
En 2001, el paleontólogo estadounidense Robert J. Gay identificó los restos de al menos tres nuevos especímenes de Dilophosaurus, este número se basa en la presencia de tres fragmentos de hueso púbico y dos fémures de tamaño diferentes en las colecciones del Museo del Norte de Arizona. Los especímenes se encontraron en 1978 en el cuadrángulo de Rock Head, a 190 kilómetros de donde se encontraron los especímenes originales, y habían sido etiquetados como un "gran terópodo". Aunque la mayoría del material está dañado, es significativo al incluir elementos no preservados en las muestras anteriores, que incluyen parte de la pelvis y varias costillas. Algunos elementos de la colección pertenecían a un espécimen infantil MNA P1.3181, el ejemplo más joven conocido de este género, y uno de los terópodos infantiles más antiguos conocidos de América del Norte, solo precedido por algunos especímenes de Coelophysis. La muestra juvenil incluye un húmero parcial, un peroné parcial y un fragmento de diente.[14] En 2005, el paleontólogo estadounidense Ronald S. Tykoski asignó un espécimen, TMM 43646-140, de Gold Spring, Arizona, a Dilophosaurus, pero en 2012 el paleontólogo estadounidense Matthew T. Carrano y sus colegas encontraron que difería en algunos detalles.[19][25]
Icnotaxones relacionados
Varios icnotaxones taxones basados en huellas fósiles se han atribuido a Dilophosaurus o terópodos similares. En 1971 Welles reportó huellas de dinosaurios de la formación Kayenta del norte de Arizona, en dos niveles a 14 metros y 112 metros por debajo de donde se encontraron los especímenes originales de Dilophosaurus. Las huellas más bajas eran tridactilas, con tres dedos, y podrían haber sido hechas por Dilophosaurus. Welles creó los nuevos icnogénero e icnoespecie, Dilophosauripus williamsi basados en ellas, nombrándolas en honor a Williams, el descubridor de los primeros esqueletos de Dilophosaurus. El espécimen tipo es un molde de yeso de una huella grande catalogado como UCMP 79690-4, con moldes de otras tres impresiones incluida sobre en él.[26] En 1984, Welles admitió que no había forma de probar o refutar que las huellas pertenecían a Dilophosaurus.[2] En 1996, los paleontólogos estadounidenses Michael Morales y Scott Bulkey informaron sobre un rastro de los icnogénero Eubrontes de la formación Kayenta formado por un gran terópodo. Notaron que podría haber sido realizado por un individuo de Dilophosaurus muy grande, pero encontraron esto poco probable, ya que estimaron que el animal que las habría hecho mediría entre 2,83 y 2,99 metros de altura a las caderas, en comparación con los 1,50 a 1, 75 metros del Dilophosaurus.[27]
.jpg.webp)
El paleontólogo polaco Gerard Gierliński examinó las huellas tridáctilas de las montañas de la Santa Cruz en Polonia y concluyó en 1991 que pertenecían a un terópodo similar al Dilophosaurus. Llamó a la nueva icnoespecie Grallator (Eubrontes) soltykovensis basada en ellos, con un molde de huella MGIW 1560.11.12 como el holotipo.[28] Huellas de una especie sin identificar de Dilophosaurus han sido halladas en Vallåkra, Suecia.[29] Algunas de las huellas fueron llevadas a museos, pero la mayor parte de estas desaparecieron en inundaciones naturales.[30] En 1994, Gierlinski y Ahlberg asignaron a estas huellas de la Formación Hoganas de Suecia a Dilophosaurus.[31] En 1994, Gierliński también asignó las huellas de la Formación Höganäs en Suecia descubierta en 1974 a G. (E.) soltykovensis.[32] En 1996, Gierliński atribuyó la pista AC 1/7 de la formación Turners Falls de Massachusetts, un rastro en reposo que creía que mostraba impresiones de plumas, de un terópodo similar al Dilophosaurus o Liliensternus, y se lo asignó al icnotaxón Grallator minisculus.[7] El espécimen AC 1/7 representa una "impresión de un dinosaurio sentado", la cual se realizó cuando un dinosaurio estaba descansando apoyando su cuerpo en el suelo, dejando una impresión de su vientre entre un par de huellas. Curiosamente, se pueden observar rastros de una cubierta similar a pelaje, la cual Gierlinski interpretó como marcas de plumas, sugiriendo que el Dilophosaurus era un dinosaurio emplumado.[31] El paleontólogo checo, Martin Kundrát, estuvo de acuerdo en que la huella mostraba impresiones de pluma en 2004, pero esta interpretación fue discutida por los paleontólogos estadounidenses Martin Lockley y colegas en 2003 y Anthony J. Martin y colegas en 2004, quienes los consideraron artefactos sedimentológicos. Martin y sus colegas también reasignaron la pista al icnotaxón Fulicopus lyellii.[8][9][10]

El paleontólogo estadounidense Robert E. Weems propuso en 2003 que las huellas de Eubrontes no fueron producidas por un terópodo sino por un sauropodomorfo similar al Plateosaurus, excluyendo al Dilophosaurus como un posible hacedor de huellas. En cambio, Weems propuso a Kayentapus hopii, otro icnotaxón nombrado por Welles en 1971, como el mejor partido para el Dilophosaurus.[33] La atribución al Dilophosaurus se basó principalmente en el gran ángulo entre las impresiones de los dígitos tres y cuatro mostradas por estas pistas, y la observación de que el pie del holotipo muestra un cuarto dígito extendido de manera similar. También en 2003, la paleontóloga estadounidense Emma Rainforth argumentó que la separación en el holotipo del pie era simplemente el resultado de la distorsión, y que Eubrontes sería un buen partido para el Dilophosaurus.[34][35]
En 2006, Weems defendió su evaluación de 2003 de Eubrontes, y propuso un animal como el Dilophosaurus como posible creador de huellas de numerosos rastros de Kayentapus de la Cantera Culpeper en Virginia. Weems sugirió que las impresiones redondeadas asociadas con algunos de estos rastros representan las impresiones de las manos que carecen de trazas de dígitos, lo que él interpretó como una traza de movimiento cuadrúpedo.[35] En 2009, los paleontólogos estadounidenses Andrew R. C. Milner y sus colegas utilizaron la nueva combinación Kayentapus soltykovensis, y sugirieron que el Dilophosauripus puede no ser distinto del Eubrontes y Kayentapus. Sugirieron que las largas marcas de garras que se utilizaron para distinguir al Dilophosauripus pueden ser un artefacto de arrastre. Descubrieron que las pistas de Gigandipus y Anchisauripus también pueden representar variaciones de Eubrontes. Señalaron que las diferencias entre icnotaxones pueden reflejar cómo el animal que realiza las pistas interactuó con el sustrato en lugar de la taxonomía. También encontraron al Dilophosaurus adecuado para una vía de huellas asignada a Eubrontes de marcas de desplazamiento y descanso catalogadas bajo el número SGDS.18.T1 del yacimiento de dinosaurios de San George en la formación Moenavede, Utah, aunque el dinosaurio en sí no se conoce en la formación, que es un poco más antigua que la formación Kayenta.[36]
Dilophosaurus "breedorum"
En 1984 Welles sugirió que el espécimen de 1964, UCMP 77270 no pertenecía a Dilophosaurus, sino a un nuevo género, basado en diferencias en el cráneo, las vértebras y la fémur. Sostuvo que ambos géneros tenían crestas, pero que la forma exacta de esta era desconocida en Dilophosaurus.[2] Welles murió en 1997, antes de que pudiera nombrar a este supuesto nuevo dinosaurio, pero la idea de que los dos eran géneros separados generalmente ha sido ignorada u olvidada desde entonces.[3] En 1999, el paleontólogo amateur americano Stephan Pickering publicó en privado el nuevo nombre Dilophosaurus "breedorum" basado en el espécimen de 1964, nombrado en honor a Breed, que había ayudado a recolectarlo. Este nombre se considera inválido ya que no cumple con los requisitos y Gay señaló en 2005 que no existen diferencias significativas entre D . "breedorum" y otros especímenes de D. wetherilli.[13][37] Carrano y sus colegas encontraron diferencias entre el espécimen de 1964 y el holotipo, pero los atribuyeron a la variación entre individuos en lugar de especies.[19]
Dilophosaurus sinensis

Un esqueleto de terópodo casi completo, KMV 8701, fue descubierto en la formación Lufeng, en la provincia de Yunnan, China, en 1987. Es similar a Dilophosaurus, con un par de crestas y un espacio que separa la premaxila del maxilar, pero difiere en algunos detalles. El paleontólogo chino Shaojin Hu lo nombró como una nueva especie de Dilophosaurus en 1993, D. sinensis, el epíteto específico proviene del griego Sinaí, refiriéndose a China.[38] En 1998, el paleontólogo estadounidense Matthew C. Lamanna y sus colegas encontraron que D. sinensis era idéntico a Sinosaurus triassicus, un terópodo de la misma formación, nombrado en 1940.[39] Esta conclusión fue confirmada por la paleontóloga china Lida Xing y sus colegas en 2013, y aunque el paleontólogo chino Guo-Fu Wang y sus colegas estuvieron de acuerdo en que la especie pertenecía a Sinosaurus en 2017, sugirieron que podría ser una especie separada, S. sinensis.[40][41]
Clasificación
Dilophosaurus ha sido examinado varias veces a través de los años y ha sido asignado a no menos de nueve grupos de terópodos. Welles en 1954 y la mayoría de los análisis filogenéticos consecutivos durante los años 1980s y 1990s clasificaron a este género como un celofisoideo grande dentro del taxón Coelophysoidea.[11][42] Sin embargo, algunos estudios más recientes han provisto diferentes grados de evidencia que implican que Dilophosaurus y varios otros "dilofosáuridos" están más cercanamente relacionados con los terópodos tetanuranos que a los verdaderos celofisoideos.[43][44]

Welles pensó a Dilophosaurus como un megalosáurido en 1954, pero revisó su opinión en 1970 después de descubrir que tenía crestas.[18][11] En 1974, Welles y el paleontólogo estadounidense Robert A. Long llegaron a la conclusión que Dilophosaurus era un ceratosauriano.[45] En 1984, Welles descubrió que Dilophosaurus presentaba características tanto de celurosauriano como de carnosaurio, los dos grupos principales en los que hasta entonces se habían dividido los terópodos, en función del tamaño corporal, y sugirió que esta división era inexacta. Encontró a Dilophosaurus como el más cercano a los terópodos que generalmente se ubicaban en la familia Halticosauridae, particularmente Liliensternus.[2]
En 1988, el paleontólogo estadounidense Gregory S. Paul clasificó a los halticosáuridos como una subfamilia de la familia Coelophysidae y sugirió que Dilophosaurus podría haber sido un descendiente directo de Coelophysis. Paul también consideró la posibilidad de que los espinosáuridos fueran dilofosáuridos supervivientes tardíos, basándose en la similitud del hocico estrecho, la posición de la fosa nasal y los dientes delgados de Baryonyx .[4] En 1994 el paleontólogo estadounidense Thomas R. Holtz colocó a Dilophosaurus en el grupo Coelophysoidea, pero separado de Coelophysidae. Colocó a Coelophysoidea en el grupo Ceratosauria.[46] En 2000, el paleontólogo estadounidense James H. Madsen y Welles dividieron Ceratosauria en las familias Ceratosauridae y Dilophosauridae, con Dilophosaurus como único miembro de esta última.[47]
Lamanna y sus colegas señalaron en 1998 que, dado que se descubrió que Dilophosaurus tenía crestas en el cráneo y se han descubierto otros terópodos de cresta similar, incluido Sinosaurus, y que esta característica no es exclusiva del género, y de uso limitado para determinar interrelaciones dentro de su grupo.[39] El paleontólogo australiano Adam M. Yates describió el nuevo género Dracovenator de Sudáfrica en 2005 y lo encontró estrechamente relacionado con Dilophosaurus y Zupaysaurus. Su análisis cladístico sugería que no pertenecían a Coelophysoidea, sino más bien a Neotheropoda, un grupo más derivado o "avanzado". Propuso que si Dilophosaurus era más derivado que Coelophysoidea, las características que compartió con este grupo pueden haber sido heredadas de terópodos más basales o "primitivos", lo que indica que los terópodos pueden haber pasado por una "etapa celofisoide" en su evolución temprana. [48]
En 2007, el paleontólogo estadounidense Nathan D. Smith y sus colegas encontraron que el terópodo crestado Cryolophosaurus era la taxon hermano de Dilophosaurus y los agruparon con Dracovenator y Sinosaurus. Este clado fue más derivado que Coelophysoidea, pero más basal que Ceratosauria, colocando los terópodos basales en una disposición tipo escalera.[49] En 2011 Sues y colegas incluyeron a Dilophosaurus en un análisis de terópodos tempranos donde aparece como taxón hermano de los terópodos jurásicose más avanzados.[50] En 2012 Carrano y sus colegas encontraron que el grupo de terópodos con cresta propuesto por Smith y sus colegas se basó en características que se relacionan con la presencia de tales crestas, pero que las características del resto del esqueleto fueron menos consistentes. En cambio, encontraron que Dilophosaurus era un celofisoide, con Cryolophosaurus y Sinosaurus siendo más derivados, como miembros basales del grupo Tetanurae.[19]
El paleontólogo belga Christophe Hendrickx y sus colegas definieron a Dilophosauridae para incluir Dilophosaurus y Dracovenator en 2015, y señalaron que si bien existe una incertidumbre general acerca de la ubicación de este grupo, parece ser un poco más derivado que Coelophysoidea y el grupo hermano de Averostra. Dilophosauridae comparte características con Coelophysoidea, como la brecha subnarial y los dientes frontales del maxilar apuntando hacia adelante, mientras que las características compartidas con Averostra incluyen una fenestra en la parte frontal del maxilar y una cantidad reducida de dientes en el maxilar superior. Sugirieron que las crestas craneales de Cryolophosaurus y Sinosaurus habían evolucionado de manera convergente, o fueron una característica heredada de un ancestro común.[51]
Filogénia
A continuación se encuentra un cladograma basado en el análisis filogenético realizado por Sues et al. en 2011, mostrando las relaciones de Dilophosaurus:[50]

| Theropoda |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
El siguiente cladograma se basa en el publicado por Hendrickx y sus colegas, basado en estudios anteriores.[51]

| Neotheropoda |
| |||||||||||||||||||||||||||||||||||||||||||||||||||
Diagnóstico
De acuerdo con Rauhut en 2000, Dilophosaurus puede ser distinguido basándose en las siguientes características, Hueso lacrimal con un borde dorso-posterior engrosado, espinas neurocervicales con una distintiva "cima" central y un "rellano" anterior y posterior, omóplatos con una expansión distal cuadrada.[52][2] Además Carrano et al. en 2012 diagnosticaron a Dilophosurus añadiendo el siguiente rasgo, crestas pares nasolacrimales extendiéndose verticalmente desde la bóveda craneana, cada una con una proyección posterior en forma de dedo.[53]
Paleobiología
Alimentación
La presencia y distribución de suturas no entrelazadas en el cráneo de algunos grupos de reptiles, incluyendo a Dilophosaurus, ha sido interpretado como evidencia de la presencia de un sistema de nivelación, impulsado por los músculos mandibulares, como una ayuda a la depredación.[54] Welles en 1984 rechazó está hipótesis e interpretó la movilidad potencial del cráneo de Dilophosaurus como signo debilidad, y afirmó que la débil conexión del premaxilar excluía la captura y sometimiento de una presa.[2] Esto llevó a la idea de que Dilophosaurus era un carroñero que consumía exclusivamente cadáveres, ya que sus dientes era demasiado frágiles como para derribar grandes presas.[15]
Welles descubrió que Dilophosaurus no tenía una mordida poderosa, debido a la debilidad causada por la brecha subnarial. Pensó que usaba sus dientes premaxilares frontales para puntear y desgarrar en lugar de morder, y los dientes superiores de más atrás para perforar y cortar. Pensó que probablemente era un carroñero en lugar de un depredador y que si mataba animales grandes, lo habría hecho con sus manos y pies en lugar de sus mandíbulas. Welles no encontró evidencia de cinesia craneal en el cráneo de Dilophosaurus, una característica que permite que los huesos individuales del cráneo se muevan en relación uno con el otro.[2]
En 1986, el paleontólogo estadounidense Robert T. Bakker descubrió que Dilophosaurus, con su enorme cuello y cráneo y sus grandes dientes superiores, había sido adaptado para matar presas grandes y lo suficientemente fuerte como para atacar a cualquier herbívoro del Jurásico Inferior.[55] En 1988, Paul descartó la idea de que Dilophosaurus era un carroñero y afirmó que los animales terrestres estrictamente carroñeros son un mito. Afirmó que el hocico de Dilophosaurus estaba mejor armado de lo que se había pensado anteriormente, y que los dientes maxilares muy grandes y delgados eran más letales que las garras. Paul sugirió que cazaba animales grandes como los prosaurópodos, y que era más capaz de matar animales pequeños que otros terópodos de un tamaño similar.[4] Paul también describió a Dilophosaurus rebotando en su cola mientras arremetía contra un enemigo, similar a un canguro.[56]
Milner y James I. Kirkland sugirieron en 2007 que Dilophosaurus tenía características que indican que pudo haber comido pescado. Señalaron que los extremos de las mandíbulas se expandieron hacia los lados, formando una "roseta" de dientes entrelazados, similar a los de los espinosáuridos, conocidos por haber comido pescado, y los gaviales , que come la mayoría de los peces. Las aberturas nasales también se retrajeron en las mandíbulas, similar a los espinosáuridos, que tienen aberturas nasales aún más retraídas, y esto puede haber limitado el agua que salpica en las fosas nasales durante la pesca. Ambos grupos también tenían brazos largos con garras bien desarrolladas, que podrían ayudar a atrapar peces. El Lago Dixie, un gran lago que se extendía desde Utah hasta Arizona y Nevada, habría proporcionado abundantes peces en el mundo "poscatalcísmico", biológicamente más empobrecido, que siguió al evento de extinción Triásico-Jurásico.[57]
Crecimiento
Welles originalmente interpretó los especímenes más pequeños de Dilophosaurus como juveniles, y el espécimen más grande como un adulto, más tarde interpretándolos como especies diferentes.[2][18] Paul sugirió que las diferencias entre los especímenes quizás se debían al dimorfismo sexual, como aparentemente también era evidente en Coelophysis, que tenía formas "robustas" y "gráciles" del mismo tamaño, que de otro modo podrían haber sido consideradas como especies separadas. Siguiendo este esquema, el espécimen más pequeño de Dilophosaurus representaría un ejemplo "gracil".[4]
En 2005 Tykoski descubrió que la mayoría de los especímenes de Dilophosaurus conocidos eran individuos juveniles, con solo el más grande adulto, según el nivel de osificación de los cartílagos de crecimiento de los huesos.[25] En 2005, Gay no encontró evidencia del dimorfismo sexual sugerido por Paul, pero supuestamente presente en Coelophysis y atribuyó la variación observada entre los especímenes de Dilophosaurus a la variación individual y ontogenia, cambios durante el crecimiento. No había dimorfismo en los esqueletos, pero no descartaba que pudiera haber habido en las crestas, se necesitaban más datos para determinar esto.[13] Sobre la base de las pequeñas crestas nasales en un ejemplar juvenil, Yates había asignado provisionalmente al género relacionado Dracovenator, sugirió que éstas habrían crecido más grande cuando el animal se convirtió en adulto.[48]
El paleontólogo estadounidense Joe S. Tkach informó sobre un estudio histológico, estudio microscópico de las características internas, de Dilophosaurus en 1996, realizado tomando secciones delgadas de huesos largos y costillas del espécimen UCMP 37303, el menos conservado de los dos esqueletos originales. Los tejidos óseos estaban bien vascularizados y tenían una estructura fibrolamelar similar a la encontrada en otros terópodos y el sauropodomorfo Massospondylus . La estructura plexiforme de los huesos sugiere un crecimiento rápido, y Dilophosaurus puede haber alcanzado una tasa de crecimiento de 30 a 35 kilogramos por año temprano en vida.[58]
Welles descubrió que los dientes de reemplazo de Dilophosaurus y otros terópodos se originaron en el interior del hueso, disminuyendo su tamaño cuanto más lejos estaban del borde alveolar. Por lo general, había dos o tres dientes de reemplazo en los alvéolos, siendo el más joven una corona pequeña y hueca . Los dientes de reemplazo emergían en el lado externo de los dientes viejos. Cuando un diente se acercó a la línea de las encías , la pared interna entre las placas interdentales se reabsorbió y formó una muesca. Cuando el diente nuevo emergió, se movió hacia fuera para centrarse en el alvéolo, y la muesca se cerró.[2]
Locomoción

Welles imaginó a Dilophosaurus como un animal activo, claramente bípedo, similar a un avestruz agrandado . Descubrió que las extremidades anteriores eran armas poderosas, fuertes y flexibles, y no se usaban para la locomoción. Notó que las manos eran capaces de agarrar y cortar, de encontrarse y llegar a dos tercios en el cuello. Propuso que en una postura sentada, el animal descansara sobre el gran "pie" de su isquion, así como su cola y sus pies.[2] En 1990 los paleontólogos estadounidenses Stephen y Sylvia Czerkas sugirieron que la pelvis débil de Dilophosaurus podría haber sido una adaptación para un estilo de vida acuático, donde el agua ayudaría a soportar su peso, y que podría haber sido un nadador eficiente. Sin embargo, encontraron dudoso que hubiera estado restringido a un medio acuoso debido a la fuerza y las proporciones de sus patas traseras, lo que lo habría hecho ágil y ágil durante la locomoción bípeda.[60]
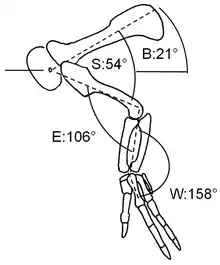
En 2005, los paleontólogos estadounidenses Phil Senter y James H. Robins examinaron el rango de movimiento en las extremidades anteriores de Dilophosaurus y otros terópodos. Descubrieron que Dilophosaurus habría podido mover su húmero hacia atrás hasta que estuvo casi paralelo a la escápula, pero no podía hacerlo hacia delante en una orientación más que vertical. El codo podría acercarse a la extensión y flexión completas en ángulo recto, pero no alcanzarlo por completo. Los dedos no parecen haber sido voluntariamente hiperextensibles, capaces de extenderse hacia atrás, más allá de su rango normal, pero pueden haber sido pasivamente hiperextensibles, para resistir la dislocación durante los movimientos violentos de las presas capturadas.[61] Un artículo de 2015 de Senter y Robins dio recomendaciones sobre cómo reconstruir la postura de la extremidad anterior en los dinosaurios bípedos, en base al examen de varios taxones, incluido Dilophosaurus. Los omóplatos se sostuvieron muy horizontalmente, la orientación de reposo del codo habría estado cerca de un ángulo recto, y la orientación de la mano no se habría desviado mucho de la del brazo inferior.[62]

Las huellas de Dilophosauripus reportadas por Welles en 1971 estaban todas en el mismo nivel, y fueron descritas como un "gallinero" de huellas de pisadas, con pocas formando un camino. Las huellas habían sido impresas en barro, lo que permitió que los pies se hundieran a una distancia de 5-10 centímetros. Las impresiones eran descuidadas, y la amplitud variable de las huellas de los dedos indica que el barro se había pegado a los pies. Las impresiones variaron de acuerdo con las diferencias en el sustrato y la manera en que se hicieron. A veces el pie se plantaba directamente, pero a menudo se producía un deslizamiento hacia atrás o hacia adelante cuando el pie bajaba. Las posiciones y los ángulos de los dedos también variaron considerablemente, lo que indica que deben haber sido bastante flexibles. Las huellas de Dilophosauripus tenían un segundo dedo desplazado con una base gruesa y garras rectas muy largas que estaban en línea con los ejes de las almohadillas para los dedos. A una de las huellas le faltaba la garra del segundo dedo del pie, tal vez debido a una lesión.[26] En 1984 Welles interpretó el hecho de que se encontraron tres individuos muy juntos, y la presencia de rastros entrecruzados cerca, como indicios de que Dilophosaurus viajó en grupos.[2] Gay estuvo de acuerdo en que pudieron haber viajado en pequeños grupos, pero notó que no había evidencia directa de esto, y que las inundaciones repentinas podrían haber recogido huesos dispersos de diferentes individuos y haberlos depositado juntos.[14]
Milner y sus colegas examinaron el camino de Utah SGDS.18.T1 en 2009, que consiste en huellas típicas con arrastres de cola y un rastro de descanso inusual, depositado en la arenisca de playa lacustre . El camino comenzó con el animal primero orientado aproximadamente en paralelo con la costa, y luego se detuvo por una berma con ambos pies en paralelo, después de lo cual bajó su cuerpo, y trajo sus metatarsianos y la callosidad alrededor de su isquion al suelo. Esto creó impresiones de "talones" simétricos e impresiones circulares del isquion. La parte de la cola más cercana al cuerpo se mantuvo fuera del suelo, mientras que el extremo más alejado del cuerpo hizo contacto con el suelo. El hecho de que el animal descansara sobre una pendiente es lo que le permitió acercar ambas manos al suelo cerca de los pies. Después de descansar, el dinosaurio se arrastró hacia adelante y dejó nuevas impresiones con sus pies, metatarsos e isquiones, pero no las manos. El pie derecho ahora pisó la huella de la mano derecha y la segunda garra del pie izquierdo hizo una marca de arrastre desde la primera posición de descanso hasta la siguiente. Después de un tiempo, el animal se puso de pie y se movió hacia delante, con el pie izquierdo primero, y una vez completamente erguido, cruzó el resto de la superficie expuesta, dejando marcas finas de arrastre con el final de la cola[36]
Agacharse es un comportamiento raramente capturado de terópodos, y SGDS.18.T1 es la única pista con impresiones inequívocas de manos de terópodo, que proporciona información valiosa sobre cómo usaron sus extremidades anteriores. Se encontró que la postura de cuclillas era muy similar a la de las aves modernas, y muestra que los primeros terópodos sostuvieron las palmas de sus manos hacia la mitad, uno hacia el otro. Como tal postura evolucionó temprano en el linaje, puede haber caracterizado a todos los terópodos. Terópodos se representan a menudo con sus palmas hacia abajo, pero los estudios de su anatomía funcional han demostrado que, como pájaros, fueron incapaces de pronación o supinación sus brazos, cruzar el radio y el cúbito de la parte inferior del brazo para girar la mano. La pista mostró que las piernas se mantuvieron simétricamente con el peso corporal distribuido entre los pies y los metatarsianos, que también es una característica que se observa en aves como las ratites . Milner y sus colegas también descartaron la idea de que la pista menor Kayentapus reportada por Weems mostrara una huella de palma hecha por un terópodo que caminaba en forma cuadrúpeda. Weems había propuesto que el animal habría podido moverse en forma cuadrúpeda cuando caminaba lentamente, mientras que los dígitos habrían sido hiperextendidos habitualmente para que solo las palmas tocaran el suelo. Milner y sus colegas descubrieron que la postura inferida es innecesaria, y sugirieron que la pista se hizo de forma similar a SGDS.18.T1, pero sin dejar rastros de los dígitos.[36]
Función de la cresta
Welles (1984) propuso que Dilophosaurus viajaba en pequeños grupos, basándose en que varios individuos se han encontrado juntos.[2] Gay (2001b) señaló que no había evidencia directa de esto y notó que “inundaciones reprentinas pudieron haber recogido el material disperso y aislado de distintos individuos y depositarlos juntos en la misma área.”[14] Se considera que las crestas del cráneo serían una ornamentación para usarse en despliegues sexuales. Esta característica tiene sentido en animales sociales o gregarios, donde haya otros miembros de la especie puedan observar e interpretar los mensajes sobre el estatus sexual.[12]

Welles admitió que las sugerencias sobre la función de las crestas de Dilophosaurus eran conjeturales, pero pensó que, aunque las crestas no tenían surcos para indicar la vascularización, podrían haberse usado para la termorregulación. También sugirió que podrían haber sido utilizados para el reconocimiento de especies u ornamentación .[2] Los Czerkas señalaron que las crestas no podrían haber sido utilizadas durante la batalla, ya que su delicada estructura se habría dañado fácilmente. Sugirieron que eran una pantalla visual para atraer a un compañero e incluso la termorregulación.[60] En 1990, el paleontólogo estadounidense Walter P. Coombs declaró que las crestas pueden haber sido mejoradas por los colores para su uso en la pantalla.[63]
En 2011, los paleontólogos estadounidenses Kevin Padian y John R. Horner propusieron que las "estructuras extrañas" en dinosaurios en general, incluyendo crestas, volantes, cuernos y cúpulas, se utilizaron principalmente para el reconocimiento de especies, y descartaron otras explicaciones como no respaldadas por evidencia. Señalaron que se conocía de muy pocos especímenes de terópodos con ornamentación craneal, incluido Dilophosaurus, para probar estadísticamente su función evolutiva y si representaban dimorfismo sexual o madurez sexual.[64]
En una respuesta a Padian y Horner el mismo año, los paleontólogos estadounidenses Rob J. Knell y Scott D. Sampson argumentaron que el reconocimiento de especies no era poco probable como una función secundaria de "estructuras extrañas" en los dinosaurios, pero que la selección sexual, utilizada en la visualización o combate para competir por compañeros, fue una explicación más probable, debido al alto costo de desarrollarlos, y porque tales estructuras parecen ser muy variables dentro de las especies.[65] En 2013 los paleontólogos británicos David E. Hone y Darren Naish criticó la "hipótesis de reconocimiento de especies" y argumentó que ningún animal existente utiliza tales estructuras principalmente para el reconocimiento de especies, y que Padian y Horner habían ignorado la posibilidad de una selección sexual mutua, donde ambos sexos están ornamentados.[66] Los estudios realizados por Gay en 2005 no muestran indicios de que hubiera dimorfismo sexual en el esqueleto de Dilophosaurus, pero sin decir nada acerca de la variación en las crestas.[13]
Paleopatología
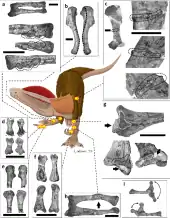
Un espécimen de Dilophosaurus wetherilli muestra daño potencial "debido a heridas o aplastamiento" en una vértebra, y un posible absceso en un húmero. Otro ejemplar de Dilophosaurus wetherilli también tenía un húmero izquierdo anormalmente pequeño comparado con un muy robusto brazo derecho, un posible ejemplo de "asimetría fluctuante". La asimetría fluctuante resulta de perturbaciones en el desarrollo y es más común en poblaciones bajo estrés y puede por tanto informar acerca de la calidad de las condiciones de vida de un dinosaurio viviendo bajo estas.[67]
Un ejemplar de Dilophosaurus del Museo de Paleontología de la Universidad de California catalogado como 'UCMP 37302 exhibe ocho diferentes patologías en su cintura escapular y los huesos de sus brazos, incluyendo tres tumores óseos, huesos rotos y luego sanados en ambos brazos, dígitos deformados y la incapacidad para utilizar uno de los miembros delanteros. Aunque no se sabe con certeza, se cree que muchas de las heridas de este espécimen fueron adquiridas durante el curso de una lucha o un accidente y que este terópodo debió haber tenido un fuerte dolor mientras se sanaba. Las patologías del animal fueron documentadas en un estudio publicado en la revista PLOS One.[68]
En un estudio de 2001 llevado a cabo por Bruce Rothschild y otros paleontólogos, se examinaron 60 huesos del pie referidos a Dilophosaurus buscando signos de fracturas por estrés, pero con resultados negativos.[69]
En 2016, Senter y Sara L. Juengst examinaron las paleopatologías del espécimen holotipo y descubrieron que presentaba el mayor y más variado número de tales enfermedades en la cintura pectoral y en la extremidad anterior de cualquier dinosaurio terópodo descrito hasta ahora, algunos de las cuales no se conocen en otro dinosaurio. Solo se conocen otros seis terópodos con más de una paleopatología en la faja pectoral y los miembros anteriores. La muestra de holotipo tenía ocho huesos afectados, mientras que no se conoce otra muestra de terópodo con más de cuatro. En su lado izquierdo tenía una escápula y radio fracturados y abscesos fibrosos en el cúbito y el hueso externo de la falange del pulgar. En el lado derecho tenía torsión de su eje humeral, tres tumores óseos en su radio, una superficie articular truncada de su tercer hueso metacarpiano y deformidades en el primer hueso de la falange del tercer dedo. Este dedo estaba permanentemente deformado e incapaz de flexionarse. Las deformidades del húmero y el tercer dedo pueden deberse a osteodisplasia, que no se había informado anteriormente de los dinosaurios, pero se conoce en las aves. Esta enfermedad, que afecta a las aves juveniles que han experimentado desnutrición, puede causar dolor en una extremidad, lo que hace que las aves prefieran usar la otra extremidad, lo que a su vez produce torsión.[70]
El número de eventos traumáticos que llevaron a estas características no es seguro, y es posible que todos hayan sido causados por un solo evento, por ejemplo al estrellarse contra un árbol o una roca durante una pelea con otro animal, lo que puede haber causado heridas punzantes. con sus garras. Como todas las heridas se habían curado, es seguro que el Dilophosaurus sobrevivió por un largo tiempo después de estos eventos, durante meses, quizás años. El uso de las extremidades anteriores para capturar presas debe haber sido comprometido durante el proceso de curación. Por lo tanto, el dinosaurio pudo haber soportado un largo período de ayuno o haber subsistido con una presa lo suficientemente pequeña como para despacharla con la boca y los pies, o con una extremidad anterior. De acuerdo con Senter y Juengst, el alto grado de dolor que el dinosaurio pudo haber experimentado en múltiples localizaciones durante largos períodos de tiempo también muestra que era un animal resistente. Señalaron que las paleopatologías en los dinosaurios no se denuncian suficientemente, y que, aunque Welles describió minuciosamente el holotipo, mencionó solo una de las patologías que encontraron.[70]
Paleoecología
Los restos de Dilophosaurus fueron recuperados en la Formación Kayenta, en el noreste de Arizona. La Formación Kayenta es parte del grupo Grupo Glen Canyon que incluye formaciones no solo del norte de Arizona sino también del sureste de Utah, el oeste de Colorado y el noroeste de Nuevo México. Esta se compone mayormente de dos facies, una dominada por deposiciones de limosas y otra dominada por areniscas. La facies de limolita se encuentra en la mayor parte de Arizona, mientras que la facies de arenisca se presenta en áras de Arizona, Utah, Colorado y Nuevo México. La formación fue depositada principalmente por ríos, siendo la facies de limolita la parte más lenta del sistema fluvial. La deposición de la Formación Kayenta fue finalizada por el campo invasor de dunas que se volvería la Arenisca Navajo.[71]
Aún no se ha realizado una datación radiométrica definitiva de la formación, y la coreelación estratigráfica disponible se ha basado en una combinación de fechas radiométricas de fósiles de vertebrados, magnetoestratigrafía y evidencia de polen.[72] Se ha conjeturado que la Formación Kayenta fue depositado durante las etapas del Sinemuriano y el Pliensbachiano del período Jurásico Inferior, es decir entre hace 196 a 183 millones de años, aproximadamente.[73]

La Formación Kayenta ha producido una pequeña pero creciente colección de organismos. La mayoría de los fósiles provienen de la facies de limonita,[74] y en general son de vertebrados. Otros organismos incluyen calizas microbianas o de "algas",[75] madera petrificada,[76] impresiones de plantas,[77] bivalvos de agua dulce y caracoles,[71] ostrácodos,[78] e icnitas de invertebrados.[75]
Los vertebrados son conocidos tanto de fósiles de partes corporales como de icnitas. Entre los vertebrados conocidos se incluyen tiburones hibodóntidos, peces óseos indeterminados, peces pulmonados, salamandras, la rana Prosalirus, la cecilia Eocaecilia, la tortuga Kayentachelys, un reptil esfenodonte, lagartos,varios crocodilomorfos primitivos incluyendo a Calsoyasuchus, Eopneumatosuchus, Kayentasuchus y Protosuchus, el pterosaurio Rhamphinion, varios terópodos como Dilophosaurus, Kayentavenator , "Syntarsus" kayentakatae, y el llamado "terópodo Shake N Bake", el sauropodomorfo Sarahsaurus, un heterodontosáurido, los dinosaurios acorazados Scelidosaurus y Scutellosaurus, los sinápsidos tritilodóntidos Dinnebiton, Kayentatherium y Oligokyphus, morganucodóntidos, el posible mamífero verdadero Dinnetherium, y un mamífero harámido. La mayoría de estos hallazgos provienen de las vecindades de Gold Spring, Arizona. Los otros fósiles de vertebrados se componen de coprolitos y las huellas de terápsidos, de animales similares a lagartos, y de varios tipos de dinosaurios.[74][75][79][76] [80][81][82][83]
Las exploraciones realizadas en las décadas de 1970 y 1980 por James M. Clark, Farish Jenkins y David E. Fastovsky y la recolección y análisis de William R. Downs han producido varios especímenes de vertebrados. En la Formación Kayenta se han hallado varios sitios de enterramientos en masa, y los restos de tres taxones de celofisoides de diferentes tamaños, los cuales representan la más diversa fauna de estos dinosaurios conocida.[84]
Tafonomía
Welles describió la tafonomía de los especímenes originales, los cambios que ocurrieron durante su decaimiento y fosilización. El esqueleto del holotipo se encontró sobre su lado derecho, y su cabeza y cuello estaban curvados hacia atrás, en la " pose de muerte " en la que a menudo se encuentran los esqueletos de los dinosaurios. Se pensaba que esta postura era opistótona, debido a espasmos mortales, en ese momento, pero en su lugar podría haber sido el resultado de cómo un cadáver estaba incrustado en los sedimentos. La espalda era recta, y las últimas vértebras dorsal se giraron en sus lados izquierdos. Las vértebras caudales se extendían irregularmente desde la pelvis, y las piernas estaban articuladas, con poco desplazamiento. Welles concluyó que los especímenes fueron enterrados en el lugar de su muerte, sin haber sido transportados mucho, pero que el holotipo parece haber sido perturbado por carroñeros, indicados por las vértebras dorsales rotadas y el cráneo aplastado.[2][85] Gay señaló que los especímenes que describió en 2001 mostraron evidencia de haber sido transportados por una corriente. Como ninguno de los especímenes está completo, es posible que hayan sido transportados a cierta distancia, o que hayan permanecido en la superficie y resistido durante algún tiempo antes del transporte. Pueden haber sido transportados por una inundación, como lo indica la variedad de animales encontrados como fragmentos y rotura de huesos.[14]
En la cultura popular

En la película Parque Jurásico, Dilophosaurus tiene una membrana retráctil en el cuello similar a la de un clamidosaurio, y tiene la capacidad de escupir un veneno cegador a los ojos para paralizar a sus víctimas. Dennis Nedry y Lewis Dodgson mueren a manos de estas criaturas, luego de cegarlos los Dilophosaurus aprovecharon para desmenuzarlos a mordidas, matándolos. No existe evidencia de que haya existido la membrana o la capacidad de escupir,[86] lo cual fue reconocido por M. Crichton como licencia literaria.[87] En la película de Steven Spielberg también se redujo el tamaño de Dilophosaurus dejándolo mucho más pequeño de lo que era en realidad. Los productos derivados de Parque Jurásico incluyeron videojuegos en los que se incluía la presencia de Dilophosaurus, como los videojuegos de simulación Jurassic Park: Operation Genesis y Jurassic World Evolution. Los videojuegos basados en las secuelas, The Lost World: Jurassic Park y Jurassic Park III, incluyeron también a Dilophosaurus. El Dilophosaurus de Parque Jurásico fue reconocido como el "único alejamiento serio de la veracidad científica" en el libro de creación de la película , y como el "más ficcionalizado" de los dinosaurios de la película en un libro sobre Stan Winston Studios, que creó el animatrónico. Para la novela, Crichton inventó la habilidad del dinosaurio para escupir veneno, explicando cómo fue capaz de matar presas, a pesar de sus fauces aparentemente débiles. El departamento de arte agregó otra característica, una capucha doblada contra su cuello que se expandió y vibró cuando el animal se preparó para atacar, similar a la del clamidosaurio. Para evitar la confusión con el Velociraptor que se muestra en la película, Dilophosaurus se presenta con solo 1,2 metros de altura, en lugar de su supuesta altura real de aproximadamente 3,0 metros. Apodado "el escupidor", el Dilophosaurus de la película se realizó a través de marionetas y requirió un cuerpo completo con tres cabezas intercambiables para producir las acciones requeridas por el guion. Las piernas separadas también se construyeron para una escena donde el dinosaurio salta. A diferencia de la mayoría de los otros dinosaurios en la película, no se emplearon imágenes generadas por computadora al mostrar el Dilophosaurus.[87][88][89]
El geólogo estadounidense J. Bret Bennington señaló que aunque Dilophosaurus no tenía un volante y no podía escupir veneno como en la película, su mordida podría haber sido venenosa, como se ha afirmado para el dragón de Komodo. Descubrió que agregar veneno al dinosaurio no era menos permisible que dar color a su piel, que también es desconocida. Si el dinosaurio tenía un volante, habría evidencia de esto en los huesos, en forma de una estructura rígida para sostener el volante, o marcas en los lugares donde los músculos utilizados para moverlos estaban unidos. También agregó que si tuviera un volante, no lo habría usado para intimidar su comida, sino más bien a un competidor, podría haber respondido a un personaje de la película que se cubría la cabeza con una capucha.[86] El propio Welles estaba "encantado" de ver a Dilophosaurus en Jurassic Park , y notó las inexactitudes pero encontró puntos menores y disfrutó de la película, y se alegró de encontrar al dinosaurio "un actor conocido internacionalmente".[90] Dennis Nedry y Lewis Dodgson murieron a manos de estas criaturas, luego de cegarlos los Dilophosaurus aprovecharon para desmenuzarlos a mordidas, matándolos.
En 2017 Dilophosaurus fue designado como el dinosaurio estatal del estado estadounidense de Connecticut, para convertirse en oficial con el nuevo presupuesto estatal en 2019. Se eligió Dilophosaurus porque se descubrieron huellas que se cree que fueron hechas por un dinosaurio relacionado en Rocky Hill en 1966, durante la excavación para la carretera interestatal 91 . Las seis pistas fueron asignadas a la icnospecie Eubrontes giganteus, que se convirtió en el fósil estatal de Connecticut en 1991. El área en la que se encontraron había sido un lago triásico y cuando se confirmó la importancia del área, la carretera se redirigió, y el área hizo un parque estatal llamado Parque Estatal Dinosaurio. En 1981, una escultura de Dilophosaurus, la primera reconstrucción a tamaño natural de este dinosaurio, fue donada al parque.[4][91][92] Dilophosaurus fue propuesto como el dinosaurio estatal de Arizona en 1998, pero los legisladores sugirieron Sonorasaurus , argumentando que Dilophosaurus no era exclusivo de Arizona. Más tarde se sugirió un compromiso que reconocería a ambos dinosaurios, pero el proyecto de ley murió cuando se reveló que los fósiles de Dilophosaurus habían sido tomados sin permiso de la Reserva Navajo.[93]
Referencias
- Xing, L.; Bell, P. R.; Rothschild, B. M.; Ran, H.; Zhang, J.; Dong, Z.; Zhang, W.; Currie, P. J. (2013). «Tooth loss and alveolar remodeling in Sinosaurus triassicus (Dinosauria: Theropoda) from the Lower Jurassic strata of the Lufeng Basin, China». Chinese Science Bulletin. doi:10.1007/s11434-013-5765-7.
- Welles, S. P. (1984). «Dilophosaurus wetherilli (Dinosauria, Theropoda), osteology and comparisons». Palaeontogr. Abt. A 185: 85-180.
- Naish, D. (2009). The Great Dinosaur Discoveries. London: A & C Black Publishers Ltd. pp. 94–95. ISBN 978-1-4081-1906-8.
- Paul, G. S. (1988). Predatory Dinosaurs of the World. New York: Simon & Schuster. pp. 258, 267–271. ISBN 978-0-671-61946-6.
- Holtz, T. R. Jr. (2012). Dinosaurs: The Most Complete, Up-to-date Encyclopedia for Dinosaur Lovers of All Ages. New York: Random House. p. 81. ISBN 978-0-375-82419-7.
- Paul, G. S. (2010). The Princeton Field Guide to Dinosaurs. Princeton: Princeton University Press. pp. 75. ISBN 978-0-691-13720-9.
- Gierliński, G. (1996). «Feather-like impressions in a theropod resting trace from the Lower Jurassic of Massachusetts». Museum of Northern Arizona Bulletin 60: 179-184.
- Kundrát, M. (2004). «When did theropods become feathered?—Evidence for pre-Archaeopteryx feathery appendages». Journal of Experimental Zoology Part B: Molecular and Developmental Evolution. 302B (4): 355-364. doi:10.1002/jez.b.20014.
- Lockley, M.; Matsukawa, M.; Jianjun, L. (2003). «Crouching theropods in taxonomic jungles: ichnological and ichnotaxonomic investigations of footprints with metatarsal and ischial impressions». Ichnos 10 (2–4): 169-177. doi:10.1080/10420940390256249.
- Martin, A. J.; Rainforth, E. C. (2004). «A theropod resting trace that is also a locomotion trace: case study of Hitchcock’s specimen AC 1/7». Geological Society of America, Abstracts with Programs 36 (2): 96. Archivado desde el original el 31 de mayo de 2004. Consultado el 2 de julio de 2018.
- Welles, S. P. (1954). «New Jurassic dinosaur from the Kayenta Formation of Arizona». Bulletin of the Geological Society of America 65 (6): 591-598. Bibcode:1954GSAB...65..591W. doi:10.1130/0016-7606(1954)65[591:NJDFTK]2.0.CO;2.
- Dodson, P. (1997) Paleoecology In: Encyclopedia of Dinosaurs, edited by Currie, P.J., and Padian, K., Academic Press.
- Carpenter, Kenneth; Gay, Robert; et al (2005). «Evidence for sexual dimorphism in the Early Jurassic theropod dinosaur, Dilophosaurus and a comparison with other related forms». The Carnivorous Dinosaurs. Indiana University Press. pp. 277–283. ISBN 0-253-34539-1.
- Gay, Robert (2001). «New specimens of Dilophosaurus wetherilli (Dinosauria: Theropoda) from the early Jurassic Kayenta Formation of northern Arizona». Western Association of Vertebrate Paleontologists annual meeting volume Mesa, Arizona 1: 1.
- Norman, David (1985). The Illustrated Encyclopedia of Dinosaurs. Nueva York: Crescent Books. pp. 62-67. ISBN 0-517-468905.
- Tykoski, R. S.; Rowe, T. (2004). «Ceratosauria». En Weishampel, D. B.; Dodson, P.; Osmolska, H., eds. The Dinosauria (2 edición). Berkeley: University of California Press. pp. 47–70. ISBN 978-0-520-24209-8.
- Welles, S. P. (1983). «Two centers of ossification in a theropod astragalus». Journal of Paleontology 57 (2): 401-401. JSTOR 1304663.
- Welles, S. P. (1970). «Dilophosaurus (Reptilia: Saurischia), a new name for a dinosaur». Journal of Paleontology 44 (5): 989. JSTOR 1302738.
- Carrano, M. T.; Benson, R. B. J.; Sampson, S. D. (2012). «The phylogeny of Tetanurae (Dinosauria: Theropoda)». Journal of Systematic Palaeontology 10 (2): 211-300. doi:10.1080/14772019.2011.630927.
- Rauhut, O. W. (2004). «The interrelationships and evolution of basal theropod dinosaurs». The Palaeontological Association 69: 213.
- Welles, S. P.; Guralnick, R. P. (1994). «Dilophosaurus Discovered». ucmp.berkeley.edu. University of California, Berkeley. Archivado desde el original el 26 de marzo de 2018. Consultado el 13 de febrero de 2018.
- Welles, Sam (2007). «Dilophosaurus Details». ucmp.berkeley.edu. University of California, Berkeley. Consultado el 17 de noviembre de 2007.
- Welles, Sam (2007). «Dilophosaurus Details». ucmp.berkeley.edu. Consultado el 17 de noviembre de 2007.
- Glut, D. F. (1997). Dinosaurs: The Encyclopedia. Jefferson: McFarland & Company, Inc. Publishers. pp. 347–350. ISBN 978-0786472222.
- Tykoski, R. S. (2005). «Anatomy, ontogeny, and phylogeny of coelophysoid theropods». UT Electronic Theses and Dissertations: 1-232.
- Welles, S.P. (1971). Dinosaur footprints from the Kayenta Formation of northern Arizona: Plateau, v. 44, pp. 27-38.
- Morales, M.; Bulkley, S. (1996). «Paleoichnological evidence for a theropod dinosaur larger than Dilophosaurus in the Lower Jurassic Kayenta Formation. The Continental Jurassic.». Museum of Northern Arizona Bulletin 60: 143-145.
- Gierliński, G. (1991). «New dinosaur ichnotaxa from the Early Jurassic of the Holy Cross Mountains, Poland». Palaeogeography, Palaeoclimatology, Palaeoecology 85 (1–2): 137-148. Bibcode:1991PPP....85..137G. doi:10.1016/0031-0182(91)90030-U.
- «Copia archivada». Archivado desde el original el 20 de octubre de 2013. Consultado el 13 de diciembre de 2012.
- http://kentlundgren.blogspot.se/2012/03/kontakt-med-trias-och-jura.html
- Glut, D. F. (1999). Dinosaurs, the Encyclopedia, Supplement 1: McFarland & Company, Inc., 442pp.
- Gierliński, G.; Ahlberg, A. (1994). «Late triassic and early jurassic dinosaur footprints in the Höganäs Formation of southern Sweden». Ichnos 3 (2): 99. doi:10.1080/10420949409386377.
- Weems, R. E. (2003). «Plateosaurus foot structure suggests a single trackmaker for Eubrontes and Gigandipus footprints». En LeTourneau, P. M.; Olsen, P. E., eds. The Great Rift Valleys of Pangea in Eastern North America 2. New York: Columbia University Press. pp. 293–313. ISBN 978-0231126762.
- Rainforth, E. C. (2003). «Revision and re-evaluation of the Early Jurassic dinosaurian ichnogenus Otozoum». Palaeontology 46 (4): 803-838. doi:10.1111/1475-4983.00320.
- Weems, R. E. (2006). «The manus print of Kayentapus minor; its bearing on the biomechanics and ichnotaxonomy of Early Mesozoic saurischian dinosaurs». New Mexico Museum of Natural History and Science Bulletin 37: 369-378.
- Milner, A. R. C.; Harris, J. D.; Lockley, M. G.; Kirkland, J. I.; Matthews, N. A.; Harpending, H. (2009). «Bird-like anatomy, posture, and behavior revealed by an Early Jurassic theropod dinosaur resting trace». PLOS One 4 (3): e4591. Bibcode:2009PLoSO...4.4591M. PMID 19259260. doi:10.1371/journal.pone.0004591.
- Mortimer, M. «Pickering's taxa 6: Dilophosaurus breedorum». Theropoddatabase.blogspot.com. Archivado desde el original el 26 de marzo de 2018. Consultado el 29 de diciembre de 2017.
- Hu, S. (1993). «A short report on the occurrence of Dilophosaurus from Jinning County, Yunnan Province». Vertebrata PalAsiatica. 1 (en chinese) (1 edición) 31: 65-69.
- Lamanna, M. C.; Holtz, T. R. Jr; Dodson, P. (1998). «A reassessment of the Chinese theropod dinosaur Dilophosaurus sinensis: Abstracts of Papers. Fifty-Eighth Annual Meeting, Society of Vertebrate Paleontology». Journal of Vertebrate Paleontology 18 (3): 57-58. JSTOR 4523942.
- Xing, L.; Bell, P. R.; Rothschild, B. M.; Ran, H.; Zhang, J.; Dong, Z.; Zhang, W.; Currie, P. J. (2013). «Tooth loss and alveolar remodeling in Sinosaurus triassicus (Dinosauria: Theropoda) from the Lower Jurassic strata of the Lufeng Basin, China». Chinese Science Bulletin 58 (16): 1931. Bibcode:2013ChSBu..58.1931X. doi:10.1007/s11434-013-5765-7.
- Guo-Fu Wang; Hai-Lu You; Shi-Gang Pan; Tao Wang (2017). «A new crested theropod dinosaur from the Early Jurassic of Yunnan Province, China». Vertebrata PalAsiatica 55 (2): 177-186.
- M. T. Carrano, R. B. J. Benson, and S. D. Sampson. 2012. The phylogeny of Tetanurae (Dinosauria: Theropoda). Journal of Systematic Palaeontology 10(2):211-300 [M. Carrano/M. Carrano]
- Yates, 2005. A new theropod dinosaur from the Early Jurassic of South Africa and its implications for the early evolution of theropods. Palaeontologia Africana. 41, 105-122.
- Smith, Makovicky, Hammer and Currie, 2007. Osteology of Cryolophosaurus ellioti (Dinosauria: Theropoda) from the Early Jurassic of Antarctica and implications for early theropod evolution. Zoological Journal of the Linnean Society. 151, 377-421.
- Welles, S. P.; Long, R. A. (1974). «The tarsus of theropod dinosaurs». Annals of the South African Museum. Annale van die Suid-Afrikaanse Museum. 64: 191-218. ISSN 0303-2515.
- Holtz Jr., T. R. (1994). «The phylogenetic position of the Tyrannosauridae: implications for theropod systematics». Journal of Paleontology 68 (5): 1100-1117. JSTOR 1306180.
- Madsen, J. H.; Welles, S. P. (2000). «Ceratosaurus (Dinosauria, Theropoda): a revised osteology». Utah Geological Survey: 1-89. Archivado desde el original el 13 de enero de 2018. Consultado el 21 de julio de 2018.
- Yates, A. M. (2005). «A new theropod dinosaur from the Early Jurassic of South Africa and its implications for the early evolution of theropods». Palaeontologia Africana 41: 105-122. ISSN 0078-8554.
- Smith, N.D., Makovicky, P.J., Pol, D., Hammer, W.R., and Currie, P.J. (2007). "The dinosaurs of the Early Jurassic Hanson Formation of the Central Transantarctic Mountains: Phylogenetic review and synthesis." In Cooper, A.K. and Raymond, C.R. et al. (eds.), Antarctica: A Keystone in a Changing World––Online Proceedings of the 10th ISAES, USGS Open-File Report 2007-1047, Short Research Paper 003, 5 p.; doi:10.3133/of2007-1047.srp003.
- Hans-Dieter Sues, Sterling J. Nesbitt, David S. Berman and Amy C. Henrici (2011). "A late-surviving basal theropod dinosaur from the latest Triassic of North America". Proceedings of the Royal Society B 278 (1723): 3459–3464
- Hendrickx, C.; Hartman, S. A.; Mateus, O. (2015). «An overview of non-avian theropod discoveries and classification». PalArch's Journal of Vertebrate Palaeontology 12 (1): 73. Archivado desde el original el 22 de junio de 2018. Consultado el 22 de julio de 2018.
- Rauhut, 2000. The interrelationships and evolution of basal theropods (Dinosauria, Saurischia). Ph.D. dissertation, Univ. Bristol [U.K.]. 440 pp.
- Carrano, Benson and Sampson, 2012. The phylogeny of Tetanurae (Dinosauria: Theropoda). Journal of Systematic Palaeontology. 10(2), 211-300.
- Bradley, O. C. 1903. The muscles of mastication and movements of the skull in Lacertilia. Zoologische Jahrbiicher, Anatomie 18: 475-486.
- Bakker, R. T. (1986). The Dinosaur Heresies. New York: William Morrow. pp. 263–264. ISBN 978-0-8217-5608-9.
- Milner, A.; Kirkland, J. (2007). «The case for fishing dinosaurs at the St. George Dinosaur Discovery Site at Johnson Farm». Survey Notes of the Utah Geological Survey 39: 1-3. Archivado desde el original el 4 de noviembre de 2017. Consultado el 23 de julio de 2018.
- Tkach, J. S. (1996). «Multi-element osteohistological analysis of Dilphosaurus wetherilli. Abstracts of Papers, Fifty-sixth Annual Meeting, Society of Vertebrate Paleontology, American Museum of Natural History, New York, New York, October 16–19». Journal of Vertebrate Paleontology 16 (3): 69. JSTOR 4523755.
- http://www.plosone.org/article/info:doi/10.1371/journal.pone.0004591
- Czerkas, S. J.; Czerkas, S. A. (1990). Dinosaurs: a Global View. Limpsfield: Dragons’ World. p. 208. ISBN 978-0-7924-5606-3.
- Senter, P.; Robins, J. H. (2005). «Range of motion in the forelimb of the theropod dinosaur Acrocanthosaurus atokensis, and implications for predatory behaviour». Journal of Zoology 266 (3): 307-318. doi:10.1017/S0952836905006989.
- Senter, P.; Robins, J. H. (2015). «Resting orientations of dinosaur scapulae and forelimbs: a numerical analysis, with implications for reconstructions and museum mounts». PLOS One 10 (12): e0144036. Bibcode:2015PLoSO..1044036S. PMID 26675035. doi:10.1371/journal.pone.0144036.
- Coombs, W. P. (1990). «Behavior patterns of dinosaurs». En Weishampel, D. B.; Osmolska, H.; Dodson, P., eds. The Dinosauria (1st edición). Berkeley: University of California Press. p. 42. ISBN 978-0-520-06727-1.
- Padian, K.; Horner, J. R. (2011). «The evolution of 'bizarre structures' in dinosaurs: biomechanics, sexual selection, social selection or species recognition?». Journal of Zoology 283 (1): 3-17. doi:10.1111/j.1469-7998.2010.00719.x.
- Knell, R. J.; Sampson, S. (2011). «Bizarre structures in dinosaurs: species recognition or sexual selection? A response to Padian and Horner». Journal of Zoology 283 (1): 18-22. doi:10.1111/j.1469-7998.2010.00758.x.
- Hone, D. W. E.; Naish, D. (2013). «The 'species recognition hypothesis' does not explain the presence and evolution of exaggerated structures in non-avialan dinosaurs». Journal of Zoology 290 (3): 172-180. doi:10.1111/jzo.12035.
- Molnar, R. E., 2001, Theropod paleopathology: a literature survey: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, p. 337-363.
- http://journals.plos.org/plosone/article?id=10.1371%2Fjournal.pone.0149140
- Rothschild, B., Tanke, D. H., and Ford, T. L., 2001, Theropod stress fractures and tendon avulsions as a clue to activity: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, p. 331-336.
- Senter, P.; Juengst, S. L.; Heymann, D. (2016). «Record-breaking pain: the largest number and variety of forelimb bone maladies in a theropod dinosaur». PLOS One 11 (2): e0149140. Bibcode:2016PLoSO..1149140S. PMID 26909701. doi:10.1371/journal.pone.0149140.
- Harshbarger, J. W.; Repenning, C. A., and Irwin, J. H. (1957). Stratigraphy of the uppermost Triassic and the Jurassic rocks of the Navajo country. Professional Paper 291. Washington, D.C.: U.S. Geological Survey.
- J. M. Clark and D. E. Fastovsky. 1986. Vertebrate biostratigraphy of the Glen Canyon Group in northern Arizona. The Beginning of the Age of the Dinosaurs: Faunal change across the Triassic-Jurassic boundary, N. C. Fraser and H.-D. Sues (eds.), Cambridge University Press 285-301
- Padian, K (1997) Glen Canyon Group In: Encyclopedia of Dinosaurs, edited by Currie, P. J., and Padian, K., Academic Press.
- Lucas, S. G.; Heckert, A. B.; Tanner, L. H. (2005). «Arizona's Jurassic fossil vertebrates and the age of the Glen Canyon Group». En Heckert, A. B.; and Lucas, S. G. editors, ed. Vertebrate paleontology in Arizona. Bulletin 29. Albuquerque, NM: New Mexico Museum of Natural History and Science. pp. 95-104.
- Luttrell, P. R., and Morales, M. 1993. Bridging the gap across Moenkopi Wash: a lithostratigraphic correlation. Aspects of Mesozoic geology and paleontology of the Colorado Plateau. Pages 111-127 in Morales, M., editor. Museum of Northern Arizona, Flagstaff, AZ. Bulletin 59.
- Jenkins, F. A., Jr. and Shubin, N. H. 1998. Prosalirus bitis and the anuran caudopelvic mechanism. Journal of Vertebrate Paleontology 18(3):495-510.
- Rigby, J. K., Hamblin, W. K., Matheny, R., and Welsh, S. L. 1971. Guidebook to the Colorado river: part 3, Moab to Hite, Utah through Canyonlands National Park. Brigham Young University Research Studies, Geology Series 18(2).
- Lucas, S. G., and Tanner L. H. 2007. Tetrapod biostratigraphy and biochronology of the Triassic-Jurassic transition on the southern Colorado Plateau, USA. Palaeogeography, Palaeoclimatology, Palaeoecology 244(1-4):242-256.
- Jenkins, F. A., Jr., Crompton, A. W., and Downs, W. R. 1983. Mesozoic mammals from Arizona: new evidence in mammalian evolution. Science 222(4629):1233-1235.
- Gay, R. 2010. Kayentavenator elysiae, a new tetanuran from the early Jurassic of Arizona. Pages 27-43 in Gay, R. Notes on early Mesozoic theropods. Lulu Press (on-demand online press).
- Curtis, K., and Padian, K. 1999. An Early Jurassic microvertebrate fauna from the Kayenta Formation of northeastern Arizona: microfaunal change across the Triassic-Jurassic boundary. PaleoBios 19(2):19-37.
- Hamblin, A. H., and Foster, J. R. 2000. Ancient animal footprints and traces in the Grand Staircase-Escalante National Monument, south-central Utah. Pages 557-568 in Sprinkel, D. A., Chidsey, T. C., Jr., and Anderson, P. B. editors. Geology of Utah’s parks and monuments. Utah Geological Association, Salt Lake City, UT. Publication 28.
- Rowe, T. B., Sues, H.-D., and Reisz, R. R. 2011. Dispersal and diversity in the earliest North American sauropodomorph dinosaurs, with a description of a new taxon. Proceedings of the Royal Society B: Biological Sciences 278(1708):1044-1053.
- Tykoski, R. S., 1998, The Osteology of Syntarsus kayentakatae and its Implications for Ceratosaurid Phylogeny: Theses, The University of Texas, December 1998.
- Reisdorf, A. G.; Wuttke, M. (2012). «Re-evaluating Moodie's opisthotonic-posture hypothesis in fossil vertebrates Part I: reptiles—the taphonomy of the bipedal dinosaurs Compsognathus longipes and Juravenator starki from the Solnhofen archipelago (Jurassic, Germany)». Palaeobiodiversity and Palaeoenvironments 92 (1): 119-168. doi:10.1007/s12549-011-0068-y.
- Bennington, J Bret (1996). «Errors in the Movie "Jurassic Park"». American Paleontologist. 4(2): 4-7.
- Crichton, Michael (1990). Jurassic Park. Alfred A. Knopf. ISBN 0-394-58816-9.
- Shay, D.; Duncan, J. (1993). The Making of Jurassic Park. New York: Boxtree Ltd. pp. 24, 35-36, 113. ISBN 978-1-85283-774-7.
- Duncan, J. (2007). The Winston Effect: The Art and History of Stan Winston Studio. London: Titan Books. pp. 177-178. ISBN 978-1-84576-150-9.
- Welles, S. P.; Guralnick, R. P. (1994). «Dilophosaurus, the actor». ucmp.berkeley.edu. University of California, Berkeley. Archivado desde el original el 26 de marzo de 2018. Consultado el 13 de febrero de 2018.
- Stone, M. (2017). «Connecticut welcomes its new state dinosaur». ctboom.com. Archivado desde el original el 25 de septiembre de 2017. Consultado el 13 de febrero de 2018.
- Rogers, O. (2016). «Discovered dinosaur tracks re-route highway and lead to state park». connecticuthistory.org. Consultado el 3 de enero de 2018.
- Gebers, S. (2018). «Arizona has no official state dinosaur, so an 11-year-old proposed one — the Sonorasaurus». azcentral. Consultado el 20 de febrero de 2018.
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
- Intro de Dilophosaurus
- Errores en Jurassic Park
- Dilophosaurus en duiops
- Dilophosaurus en siplemano
- Dilophosaurus dentro de Neotherapoda en The Thescelsosaurus (en inglés)
- Significado y pronunciación del nombre (en inglés)
- Dilophosaurus wetherilli en The Theropod Database (en inglés)
| Control de autoridades |
|
|---|
 Datos: Q271710
Datos: Q271710 Multimedia: Dilophosaurus / Q271710
Multimedia: Dilophosaurus / Q271710 Especies: Dilophosaurus
Especies: Dilophosaurus