Eukaryota
En biología y taxonomía, Eukaryota o Eukarya (del griego: εὖ eu —‘bueno’, ‘bien’, 'verdadero'— y κάρυον karyon —‘nuez’, ‘carozo’, ‘núcleo’—) es el dominio (o imperio) que incluye los organismos formados por células con núcleo verdadero. La castellanización adecuada del término es eucariota o eucarionte.[5] Estos organismos constan de una o más células eucariotas, abarcando desde organismos unicelulares hasta verdaderos pluricelulares en los que las diferentes células se especializan para diferentes tareas y que, en general, no pueden sobrevivir de forma aislada.
| Eucariota | ||
|---|---|---|
| Rango temporal: 2200–0Ma Paleoproterozoico – Reciente | ||
 | ||
| Taxonomía | ||
| Dominio: |
Eukaryota Chatton 1925, orth. em. Fott 1971 o Eukarya Margulis & Chapman 2009 | |
| Reinos y supergrupos | ||
Clasificación taxonómica:[1] En otros sistemas, en lugar de Protista va:[2] Clasificación filogenética[3][4]
| ||
| Sinonimia | ||
| ||
Pertenecen al dominio o imperio eucariota los reinos de los animales, plantas y hongos, así como varios grupos incluidos en el parafilético reino Protista. Todos ellos presentan semejanzas a nivel molecular (estructura de los lípidos, proteínas y genoma), comparten un origen común, y principalmente, comparten el plan corporal de los eucariotas, muy diferente al de procariotas.
Con excepción de algunos organismos unicelulares, el ciclo de vida eucariota alterna una fase haplonte y otra diplonte, que se consigue mediante la alternancia de meiosis y fecundación, procesos que dan células haplontes y diplontes respectivamente.
Estructura celular

Las células eucariotas son generalmente mucho más grandes que las procariotas y están mucho más compartimentadas. Poseen una gran variedad de membranas con núcleo rodeado de la envoltura nuclear, retículo endoplasmático y aparato de Golgi, además de mecanismos para la gemación y fusión de vesículas, incluida la exocitosis y endocitosis. Estructuras internas llamadas orgánulos se encargan de realizar funciones especializadas dentro de la célula. Presencia de lisosomas, peroxisomas y mitocondrias.
También caracteriza a todos los eucariotas un esqueleto interno o endoesqueleto, en este caso llamado citoesqueleto, formado por dos entramados de proteínas: el sistema de microtúbulos y el sistema contráctil de actina/miosina, que desempeñan un papel importante en la definición de la organización y forma de la célula, en el tráfico intracelular (por ejemplo, los movimientos de vesículas y orgánulos) y en la división celular. El característico flagelo eucariota y sus motores moleculares asociados se encuentran anclados al citoesqueleto.
El ADN de las células eucariotas está contenido en un núcleo celular separado del resto de la célula por una doble membrana permeable. El material genético se divide en varios bloques lineales llamados cromosomas, que son separados por un huso microtubular durante la división nuclear. Los cromosomas contienen histonas, varios replicones, centrómeros y telómeros. Hay un característico ciclo celular con segregación mitótica y reproducción sexual por meiosis. Se incluye un complejo de poros nucleares, transporte trans-membranal de ARN y proteínas a través de la envoltura nuclear, intrones y nuevos patrones de procesamiento del ARN utilizando espliceosomas.
Para una comparación con las características procariotas, véase: Tabla comparativa.

Plan corporal
La célula eucariota debe en gran parte su forma y capacidad de movimiento al citoesqueleto, ya que le otorga rigidez y flexibilidad. En los organismos flagelados ancla los flagelos al resto de la célula y permite su batido durante la locomoción o para la creación de corrientes de agua que le lleven el alimento. En los organismos ameboides permite la extensión de "pies" o seudópodos para la locomoción o la alimentación. También fija los surcos de alimentación de los excavados y el complejo apical que permite a los apicomplejos entrar en las células parasitadas.
Solo después de desarrollar su citoesqueleto pudo el eucariota ancestral realizar la fagocitosis, ya que es este el que, mediante crecimiento diferencial de sus fibras, logra que la célula se deforme para que la fagocitosis ocurra. La fagocitosis es también una propiedad ancestral de los eucariotas, si bien se ha perdido en grupos que se adaptaron a otras formas de alimentación. Hongos y plantas perdieron esta capacidad al desarrollar una pared celular rígida externa a la célula, pero ya contaban con otros modos de nutrición, la saprotrofia o el parasitismo en hongos y la fotosíntesis en plantas.
La mitocondria, derivada de la fagocitosis y posterior simbiogénesis de una proteobacteria, permitió al eucariota ancestral la respiración aerobia y con ello aprovechar al máximo la energía contenida en la materia orgánica. Como no es sorprendente en la evolución de un carácter tan antiguo, en varios grupos la mitocondria ha perdido esa capacidad ancestral y a cambio se ha modificado para cumplir otras funciones. También proceden de un evento de endosimbiosis los cloroplastos, en este caso con una cianobacteria, que permiten a las plantas realizar la fotosíntesis. Posteriormente otros grupos de eucariotas consiguieron sus cloroplastos mediante la endosimbiosis secundaria con un alga verde o roja.
Reproducción

Además de la división asexual de las células (mitosis), la mayoría de los eucariontes tiene algún proceso de reproducción sexual basado en la meiosis que no se encuentra entre los procariontes. La reproducción de los eucariontes típicamente implica la existencia de una fase haploide, donde está presente solamente una copia de cada cromosoma en las células, y diploide, donde están presentes dos. Las células diploides surgen por fusión nuclear (fecundación) y las haploides, por meiosis. En los organismos multicelulares, se distinguen tres tipos de ciclos biológicos:
- Ciclo haplonte: los organismos que presentan este ciclo son haploides durante su fase adulta. El cigoto es diploide y la meiosis tiene lugar tras la fecundación.
- Ciclo diplonte: los individuos maduros son diploides y forman gametos haploides por meiosis, que se fusionan para dar lugar a un nuevo organismo diploide.
- Ciclo haplodiplonte: se produce alternancia de generaciones entre individuos haploides y diploides.
Los organismos unicelulares pueden reproducirse asexualmente por bipartición, gemación o esporulación y sexualmente mediante gametos o por conjugación.[6]
En los eucariontes, la relación de superficie frente a volumen es más pequeña que los procariontes, y así tienen tasas metabólicas más bajas y tiempos de generación más largos.
Origen
El origen de la célula eucariota es el proceso biológico más revolucionario desde el origen de la vida desde varios puntos de vista, como es el caso de la morfología, desarrollo evolutivo, estructura genética, relaciones simbióticas y ecología.[7] Todas las células complejas son de este tipo y constituyen la base de casi todos los organismos pluricelulares.[8] Aunque no hay acuerdo sobre cuándo se originaron los eucariotas, en general, se ha sugerido a comienzos del Paleoproterozoico hace unos 2500 millones de años.[9][10] Hasta ahora el fósil más antiguo que puede considerarse eucariota tiene 2200 millones de años y se le conoce como Diskagma el cual representa los primeros indicios de vida pluricelular junto con la biota francevillense de hace 2100 millones de años. La separación entre los eucariotas y su grupo hermano las arqueas Asgard se estimó a finales del Arcaico (periodo Neoarcaico).[10]
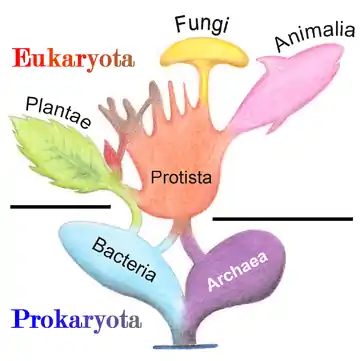
Eukarya se relaciona con Archaea desde el punto de vista del ADN nuclear y de la maquinaria genética, y ambos grupos son clasificados a veces juntos en el clado Neomura. Desde otros puntos de vista, tales como por la composición de la membrana, se asemejan más a Bacteria. Se han propuesto para ello tres posibles explicaciones principales:[12][13][14]
- Los eucariontes resultaron de la fusión completa de dos o más células, el citoplasma procedente de una bacteria y el núcleo de una arquea.
- Los eucariontes se desarrollaron de las arqueas y adquirieron sus características bacterianas a partir de las proto-mitocondrias.
- Los eucariontes y las arqueas se desarrollaron independientemente a partir de una bacteria modificada.
Cada vez son mayores las evidencias que parecen demostrar que el origen eucariota es producto de la fusión de una arquea y una bacteria. Mientras el núcleo celular tiene elementos genéticos relacionados con las arqueas, las mitocondrias y la membrana celular tienen características bacterianas. La fusión genética es más evidente al constatar que los genes informativos parecen de origen arqueano y los genes operacionales de origen bacteriano. En todo caso también es cierto que un cierto número de rasgos presentes exclusivamente en los eucariontes son difíciles de explicar por medio de un evento de fusión.[15]
Tampoco está claro el retraso de mil millones de años entre el origen de los eucariotas y su diversificación, pues las bacterias dominaron la biosfera hasta hace unos 800 millones de años.[16] Este intervalo de estabilidad ambiental, litosférica y evolutiva se conoce con el nombre de "boring billion" ("millardo -de años- aburrido").[17] Este retraso quizá se debiera simplemente a la dificultad de introducción en una biosfera ocupada enteramente por los procariotas. Otras explicaciones están relacionadas con la lenta evolución de los eucariotas o con el aumento del oxígeno, que no alcanzó los niveles ideales para los eucariotas hasta el final de dicho período. El diseño compartimentado de la célula eucariota es el más adecuado para el metabolismo aerobio y es comúnmente aceptado que todos los eucariotas actuales, incluidos los anaerobios, descienden de antecesores aerobios con mitocondrias.[18][19]
Evolución

Algún tiempo después de que surgiera la primera célula eucariota se produjo una radiación explosiva que las llevó a ocupar la mayoría de los nichos ecológicos disponibles.
Evolución unicelular
La primera célula eucariota era probablemente flagelada, aunque con tendencias ameboides al no tener una cubierta rígida.[21] Desde el antecesor flagelado, algunos grupos perdieron ulteriormente los flagelos, mientras que otros se convirtieron en multiflagelados o ciliados. Cilios y flagelos (incluidos los que tienen los espermatozoides) son estructuras homólogas con nueve dobletes de microtúbulos que se originan a partir de los centriolos.[22]
El carácter ameboide surgió varias veces a lo largo de la evolución de los protistas dando lugar a los diversos tipos de seudópodos de los distintos grupos. El que los ameboides procedan de los flagelados y no al revés, como se pensaba en el pasado, tiene como base estudios moleculares (fusión, partición o duplicación de genes, inserción o borrado de intrones, etc.).[23] [24]
Está generalmente aceptado que los cloroplastos se originaron por endosimbiosis de una cianobacteria y que todas las algas eucariotas evolucionaron en última instancia de antepasados heterótrofos. Se piensa que la diversificación primaria de la célula eucariota tuvo lugar entre los zooflagelados: células predadoras no fotosintéticas con uno o más flagelos para nadar, y a menudo también para generar corrientes de agua con las que capturar a las presas.[21]
En la actualidad hay discrepancia en dónde debe ponerse la raíz del árbol de Eukarya. La posibilidad más aceptada es situarlo entre o próximo a los excavados, que serían el grupo basal de los eucariontes.[3][25]
Evolución pluricelular

Durante la primera parte de su historia los eucariontes permanecieron unicelulares. A partir del período Ediacárico los pluricelulares comienzan a profilerar, aunque el proceso con seguridad comenzó bastante antes. Los organismos unicelulares de vida colonial comenzaron a cumplir funciones específicas en una zona del colectivo. Se formaron así los primeros tejidos y órganos. La pluricelularidad se desarrolló independientemente en varios grupos de eucariontes: plantas, hongos, animales, algas rojas, algas pardas y mohos mucilaginosos. A pesar de su pluricelularidad, estos dos últimos grupos se siguen clasificando en el reino Protista.
Las algas verdes, las primeras plantas, se desarrollaron para formar las primeras hojas. En el Silúrico surgen las primeras plantas terrestres y de ellas las plantas vasculares o cormófitas.
Los hongos unicelulares constituyeron filas de células o hifas que agrupadas se convirtieron en organismos pluricelulares absortivos con un marcado micelio. Inicialmente, los hongos fueron acuáticos y probablemente en el período Silúrico apareció el primer hongo terrestre, justo después de la aparición de las primeras plantas terrestres. Estudios moleculares sugieren que los hongos están más relacionados con los animales que con las plantas.
El reino animal comenzó con organismos similares a los actuales poríferos que carecen de verdaderos tejidos. Posteriormente se diversifican para dar lugar a los distintos grupos de invertebrados y vertebrados.
Clasificación y filogenia
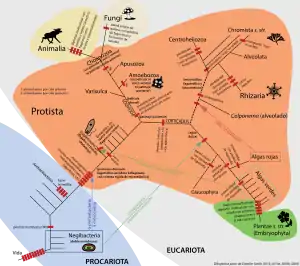

Los eucariontes se dividen tradicionalmente en cuatro reinos: Protista, Plantae, Animalia y Fungi (aunque Cavalier-Smith 2004,[30] 2015[31] reemplaza Protista por dos nuevos reinos, Protozoa y Chromista). Esta clasificación es el punto de vista generalmente aceptado en actualidad, aunque ha de tenerse en cuenta que el reino Protista, definido como los eucariontes que no encajan en ninguno de los otros tres grupos, es parafilético. Por esta razón, la diversidad de los protistas coincide con la diversidad fundamental de los eucariontes.
La reciente clasificación de Adl et al. (2018)[3] evita la clasificación en reinos, sustituyéndola por una acorde con la filogenia actualmente conocida, en la que por otra parte a los clados o taxones no se les atribuye ya categoría alguna, para evitar los inconvenientes que suponen éstas para su posterior actualización. Los principales grupos de esta clasificación (equivalentes a reinos en clasificaciones anteriores) es como sigue:
- Archaeplastida (= Primoplantae). Incluye plantas, algas verdes, algas rojas y glaucofitas. Estos organismos fotosintéticos obtuvieron sus cloroplastos por endosimbiosis primaria de una cianobacteria.
- Sar (Stramenopiles, Alveolata y Rhizaria). Este clado ancestralmente pudiera ser fotosintético por endosimbiosis secundaria de un alga roja. Cavalier-Smith lo incluye en el reino Chromista.
- Stramenopiles (= Heterokonta). Incluye algas pardas, diatomeas, oomicetos, etc.
- Alveolata. Incluye ciliados, apicomplejos y dinoflagelados.
- Rhizaria. Incluye a foraminíferos, radiolarios y varios ameboflagelados.
- Amoebozoa. Algunas amebas y hongos mucosos.
- Opisthokonta. Incluye animales, hongos y coanozoos.
- Excavata. Diversos flagelados heterótrofos o fotosínteticos, estos últimos por endosimbiosis secundaria de un alga verde. Este grupo es probablemente parafilético, por lo que debería dividirse en Metamonada, Discoba y Malawimonada.
Algunos grupos de protistas tienen una clasificación dudosa, en particular Cryptista (criptofitas) y Haptista (haptofitas), mientras que otros parecen situarse fuera de los grandes grupos, en particular CRuMs, Ancyromonadida y Hemimastigophora.
Adicionalmente se reconocen dos agrupaciones más grandes. Diaphoretickes (o Corticata) engloba a Archaeplastida y Sar, mientras que Amorphea (o Podiata) agrupa a Amoebozoa y Opisthokonta. Nótese que una forma ameboide o flagelar no indica la pertenencia a un grupo taxonómico concreto, como se creía en clasificaciones tradicionales, creando grupos artificiales desde el punto de visto evolutivo (ver polifilia).
El siguiente árbol filogenético muestras las relaciones entre los principales grupos de eucariotas según algunos estudios filogenéticos recientes:[32][33][34][35][36][37][38][39]
| Eukaryota |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||






Véase también
Referencias
- Whittaker, R. H. (1969). «New concepts of kingdoms of organisms». Science 163: 150-160.
- Cavalier-Smith, T. (1998). «A revised six-kingdom system of life». Biological Reviews of the Cambridge Philosophical Society (Cambridge University Press) 73: 203-266. doi:10.1017/S0006323198005167
- Adl, Sina M., David Bass, Christopher E. Lane, Julius Lukeš, Conrad L. Schoch, Alexey Smirnov, Sabine Agatha et al. Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes. Journal of Eukaryotic Microbiology (2018).
- Thomas Cavalier-Smith et al. 2015, Multiple origins of Heliozoa from flagellate ancestors: New cryptist subphylum Corbihelia, superclass Corbistoma, and monophyly of Haptista, Cryptista, Hacrobia and Chromista. Molecular Phylogenetics and Evolution Volume 93, December 2015, Pages 331–362
- Real Academia Española. «eucariota». Diccionario de la lengua española (23.ª edición). Consultado el 15 de julio de 2015.
- «Métodos de reproducción en procariotas y eucariotas unicelulares». Hiru. Departamento de Educación, Política Lingüística y Cultura del Gobierno Vasco.
- Lynn Margulis & Michael J Chapman, 1982-1998-2009, "Kingdoms and Domains: An Illustrated Guide to the Phyla of Life on Earth." p53
- W. Martin & M.J. Russell (1992). «On the origins of cells: a hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells». Philosophical Transactions of the Royal Society B.
- Jürgen F. H. Strassert, Iker Irisarri, Tom A. Williams & Fabien Burki (2021). A molecular timescale for eukaryote evolution with implications for the origin of red algal-derived plastids.
- Emilie Neveu, Dany Khalifeh, Nicolas Salamin, Dirk Fasshauer, (2020). Prototypic SNARE Proteins Are Encoded in the Genomes of Heimdallarchaeota, Potentially Bridging the Gap between the Prokaryotes and Eukaryotes. Current Biology.
- M. Rivera & J. Lake 2004, The ring of life provides evidence for a genome fusion origin of eukaryotes Nature 431, 152-155
- S. L. Baldauf (2003). «The Deep Roots of Eukaryotes». Science 300 (5626): 1703-1706. doi 10.1126/science.1085544.
- T.M. Embley and W. Martin (2006), Eukaryotic evolution, changes and challenges, Nature Reviews, Vol 440:30, March 2006, doi:10.1038/nature04546
- Poole A, Penny D (2007). «Evaluating hypotheses for the origin of eukaryotes». Bioessays 29 (1): 74-84. PMID 17187354.
- Poole, A. 2009. Mi nombre es LUCA – El último Ancestro Universal Común. Archivado el 16 de enero de 2014 en Wayback Machine. Un artículo original de ActionBioscience
- Butterfield, N. J. (2007). Macroevolution and macroecology through deep time. Palaeontology, 50(1), 41-55.
- Boenigk, J.; Wodniok, S.; Glücksman, E. (2015). Biodiversity and Earth History. Springer. pp. 58-59. ISBN 978-3-662-46394-9.
- Cavalier-Smith, T. (2006), Rooting the tree of life by transition analyses, Biol Direct. 1: 19. doi: 10.1186/1745-6150-1-19.
- Pisani D, Cotton JA, McInerney JO (2007). «Supertrees disentangle the chimerical origin of eukaryotic genomes». Mol Biol Evol. 24 (8): 1752-60. PMID 17504772.
- Biocyclopedia 2012, Endosymbiosis and Origin of Eukaryotic Algae
- Cavalier-Smith, T. (2006) Protozoa: the most abundant predators on earth Archivado el 15 de septiembre de 2015 en Wayback Machine.,Microbiology Today, Nov. 2006, pp. 166-167.
- D.R. Mitchell (2006), The Evolution of Eukaryotic Cilia and Flagella as Motile and Sensory Organelles en Eukaryotic Membranes and Cytoskeleton: Origins and Evolution, editado por Gáspár Jékely, Eurekah.com, ISBN 978-0-387-74020-1.
- Cavalier-Smith, T. (2003). «Protist phylogeny and the high-level classification of Protozoa». European Journal of Protistology 39 (4): 338-348.
- A. Stechmann, T. Cavalier-Smith (2003), The root of the eukaryote tree pinpointed, Current Biology, Vol. 13, No. 17, pp. R665-R666.
- Cavalier-Smith, T. (2013). Early evolution of eukaryote feeding modes, cell structural diversity, and classification of the protozoan phyla Loukozoa, Sulcozoa, and Choanozoa. European journal of protistology, 49(2), 115-178.
- Cavalier-Smith, T. (2010). Deep phylogeny, ancestral groups and the four ages of life. Philosophical Transactions of the Royal Society B: Biological Sciences, 365(1537), 111-132.
- Cavalier-Smith, T. (1998). A revised six-kingdom system of life Archivado el 4 de marzo de 2016 en Wayback Machine.. Biological Reviews, 73(3), 203-266.
- Cavalier-Smith, T. (2010). Kingdoms Protozoa and Chromista and the eozoan root of the eukaryotic tree. Biology Letters, 6(3), 342-345.
- Cavalier-Smith, T. (2009). Megaphylogeny, Cell Body Plans, Adaptive Zones: Causes and Timing of Eukaryote Basal Radiations. Journal of eukaryotic microbiology, 56(1), 26-33.
- Cavalier-Smith, T. (2004), "Only six kingdoms of life", Proceedings: Biological Sciences, Volume 271, Number 1545/June 22, pp. 1251-1262. DOI 10.1098/rspb.2004.2705
- Ruggiero, M. A., Gordon, D. P., Orrell, T. M., Bailly, N., Bourgoin, T., Brusca, R. C., Cavalier-Smith, T., Guiry, M.D. y Kirk, P. M. (2015). A Higher Level Classification of All Living Organisms.
- Romain Derelle et al 2015, Bacterial proteins pinpoint a single eukaryotic root. Proceedings of the National Academy of Sciences 112(7) · January 2015 with 248 Reads DOI: 10.1073/pnas.1420657112
- Thomas Cavalier-Smith et al 2015, Multigene phylogeny resolves deep branching of Amoebozoa. Molecular Phylogenetics and Evolution Volume 83, February 2015, Pages 293–304
- Fabien Burki et al. 2015, Untangling the early diversification of eukaryotes: a phylogenomic study of the evolutionary origins of Centrohelida, Haptophyta and Cryptista. The Royal Society 2016 DOI: 10.1098/rspb.2015.2802
- Brown, M. W., Heiss, A. A., Kamikawa, R., Inagaki, Y., Yabuki, A., Tice, A. K., ... & Roger, A. J. (2018). Phylogenomics places orphan protistan lineages in a novel eukaryotic super-group. Genome biology and evolution, 10(2), 427-433.
- Lax, Gordon; Eglit, Yana; Eme, Laura; Bertrand, Erin M.; Roger, Andrew J.; Simpson, Alastair G. B. (2018-11-14). "Hemimastigophora is a novel supra-kingdom-level lineage of eukaryotes". Nature. 564 (7736): 410–414. Bibcode:2018Natur.564..410L. doi:10.1038/s41586-018-0708-8. ISSN 0028-0836. PMID 30429611
- Tikhonenkov, Denis V.; Mikhailov, Kirill V.; Gawryluk, Ryan M. R.; Belyaev, Artem O.; Mathur, Varsha; Karpov, Sergey A.; Zagumyonnyi, Dmitry G.; Borodina, Anastasia S.; Prokina, Kristina I.; Mylnikov, Alexander P.; Aleoshin, Vladimir V.; Keeling, Patrick J. (2022). «Microbial predators form a new supergroup of eukaryotes». Nature 612 (7941): 714-719. Bibcode:2022Natur.612..714T. PMID 36477531. S2CID 254436650. doi:10.1038/s41586-022-05511-5.
- Schön, Max E.; Zlatogursky, Vasily V.; Singh, Rohan P.; Poirier, Camille; Wilken, Susanne; Mathur, Varsha; Strassert, Jürgen F. H.; Pinhassi, Jarone; Worden, Alexandra Z.; Keeling, Patrick J.; Ettema, Thijs J. G.; Wideman, Jeremy G.; Burki, Fabien (17 de noviembre de 2021). «Single cell genomics reveals plastid-lacking Picozoa are close relatives of red algae». Nature Communications 12 (1): 6651. PMC 8599508. PMID 34789758. doi:10.1038/s41467-021-26918-0.
- Cavalier-Smith T (2022). «Ciliary transition zone evolution and the root of the eukaryote tree: implications for opisthokont origin and classification of kingdoms Protozoa, Plantae, and Fungi». Protoplasma 259: 487-593. PMC 9010356. PMID 34940909. doi:10.1007/s00709-021-01665-7.
Enlaces externos
 Wikispecies tiene un artículo sobre Eukaryota.
Wikispecies tiene un artículo sobre Eukaryota. Wikimedia Commons alberga una categoría multimedia sobre Eukaryota.
Wikimedia Commons alberga una categoría multimedia sobre Eukaryota.
| Control de autoridades |
|
|---|