Historia de la vida
La historia de la vida en la Tierra cuenta los procesos por los cuales los organismos vivos han evolucionado, desde el origen de la vida en la Tierra, hace unos 4400 millones de años[1] hasta la gran diversidad presente en los organismos actuales. De la misma forma trata sobre cómo los aspectos ambientales, en forma de catástrofes globales, cambios climáticos o uniones y separaciones de continentes y océanos, han condicionado su desarrollo. Las similitudes entre todos los organismos actuales indican la existencia de un ancestro común universal del cual todas las especies conocidas han divergido a través de procesos evolutivos.[2]
Antecedentes
Tras el Big Bang el universo no reunía las condiciones para que se formara la vida, para ello fue necesario que pasara cierto tiempo y sucedieran ciertos fenómenos. En los primeros momentos del universo los únicos elementos existentes eran hidrógeno y helio, pero la vida requiere una gran diversidad de elementos. Estos primeros elementos se acumularon formando estrellas, las cuales tras su colapso como supernova dieron lugar al resto de elementos necesarios para que se formen las biomoléculas. Creados estos elementos, la vida seguía sin ser posible hasta que se formaran planetas por acreción que contuvieran dicha diversidad de elementos. El tiempo mínimo para que sucedan estos fenómenos -es decir para que acabe el ciclo de una generación de estrellas y tras ello se formen planetas con los restos de la supernova- es no menos de 4000 millones de años.[3]
Los organismos dominantes de la vida en el Arcaico temprano fueron bacterias y arqueas, que coexistieron formando alfombras microbianas (también conocidas como «tapetes» o «esteras microbianas») y estromatolitos. Muchos de los sucesos más importantes en la evolución temprana, se cree, han tenido lugar dentro de ellos.[4] La evolución de la fotosíntesis oxigénica, alrededor de hace 3500 millones de años, condujo a la oxigenación de la atmósfera, que comenzó hace alrededor de 2400 millones de años.[5] La evidencia más temprana de eucariotas (células complejas con organelos), data de hace 2200 millones de años,[6] y si bien pudieron haber estado presente antes, su diversificación acelerada comenzó cuando empezaron a utilizar el oxígeno en su metabolismo. Más tarde, alrededor de hace 1700 millones de años, los organismos multicelulares comenzaron a aparecer, con la diferenciación celular cada célula del organismo comenzó realizar funciones especializadas.[7]
Las primeras plantas terrestres datan de alrededor de 450 millones de años,[8] aunque la evidencia sugiere que la espuma de algas se formó en la tierra tan pronto como hace 1200 millones de años. Las plantas terrestres tuvieron tanto éxito que se cree que han contribuido a la extinción del Devónico tardío.[9] Los animales invertebrados aparecen durante el período Ediacárico,[10] mientras que los vertebrados se originaron hace alrededor de 525 millones de años durante la explosión cámbrica.[11]
Durante el período Pérmico, los sinápsidos, entre los que se encontraban los ancestros de los mamíferos, dominaron la tierra[12] pero el evento de extinción del Pérmico-Triásico hace 251 millones de años estuvo a punto de aniquilar toda la vida compleja.[13] Durante la recuperación de esta catástrofe, los arcosaurios se convirtieron en los vertebrados terrestres más abundantes, desplazando a los terápsidos a mediados del Triásico.[14] Un grupo de arcosaurios, los dinosaurios, dominaron los períodos Jurásico y Cretácico,[15] con los antepasados de los mamíferos sobreviviendo sólo como pequeños insectívoros.[16] Después de la extinción masiva del Cretácico-Terciario hace 65 millones de años que acabó con los dinosaurios no aviarios[17] los mamíferos aumentaron rápidamente en tamaño y diversidad.[18] Estas extinciones masivas pudieron haber acelerado la evolución, proporcionando oportunidades para que nuevos grupos de organismos se diversifiquen.[19]
La evidencia fósil y la paleobiología indican que las plantas con flores aparecieron y se diversificaron rápidamente en el Cretácico temprano, entre hace 130 millones de años y 90 millones de años, probablemente ayudado por coevolución con los insectos polinizadores. Las plantas con flores y el fitoplancton marino siguen siendo los principales productores de materia orgánica. Los insectos sociales aparecieron alrededor del mismo tiempo que las plantas con flores. A pesar de que ocupan sólo una pequeña parte del "árbol genealógico" de insectos, ahora forman más de la mitad de la masa total de los insectos. Los seres humanos evolucionaron de un linaje de los primeros hominoideos erguidos cuyos fósiles datan de más de 6 millones de años. A pesar de que los primeros miembros de este linaje tenían cerebros del tamaño de chimpancés, hay signos de un aumento constante en el tamaño del cerebro después de unos 3 millones de años.
Historia temprana de la Tierra
Tabla geológica de eventos principales
Notas
- a) Los eones Hádico, Arcaico y Proterozoico se agrupan en el Tiempo Precámbrico, también denominado Criptozoico.
- b) Todas las fechas se dan en millones de años para el inicio de la época en cuestión. Las fechas marcadas con
 son aproximadas y están basadas en acuerdos internacionales con GSSP. Las fechas del Precámbrico, excepto el inicio del periodo Ediacárico, se han fijado cronométricamente por convenio.
son aproximadas y están basadas en acuerdos internacionales con GSSP. Las fechas del Precámbrico, excepto el inicio del periodo Ediacárico, se han fijado cronométricamente por convenio. - c) Una reciente propuesta de la ICS pretendía eliminar el Cuaternario de la nomenclatura y extender el Neógeno hasta el presente, pero la propuesta no prosperó.
- d) En América del Norte, el Carbonífero se subdivide en los períodos Misisípico y Pensilvánico.
- e) Descubrimientos hechos durante el último cuarto de siglo XX han cambiado substancialmente la forma de ver los eventos geológicos y paleontológicos inmediatamente anteriores al Cámbrico. La nomenclatura no se ha estabilizado. El término Neoproterozoico es utilizado aquí, pero otros escritores podrían igualmente usar otros términos como 'Ediacariano', 'Vendiano', 'Varangiano', 'Precámbrico', 'Protocambriano', 'Eocambriano', o podrían haber extendido el período de duración del Cámbrico. Todos estos términos son considerados como un subconjunto del Proterozoico más que como un período entre el Proterozoico y el Paleozoico.
- f) La Comisión Internacional de Estratigrafía (ICS), (Véase: Global Boundary Stratotype Section and Point (GSSP) of the International Commission of Stratigraphy (status 2012-2016), e International Stratigraphic Chart, 2016) no reconoce subdivisiones en el eón Hádico, pero existe una propuesta que se inspira en la escala de tiempo geológico lunar que la subdivide en las eras Ímbrico, Nectárico, Grupos Basin y Críptico (W. Harland, R. Armstrong, A. Cox, L. Craig, A. Smith, D. Smith (1990). A Geologic time scale 1989. Cambridge University Press.).
Introducción
Los fragmentos más antiguos de meteoritos encontrados en la Tierra tienen aproximadamente 4540 millones de años; esto, asociado principalmente con la datación de antiguos yacimientos de plomo, ha hecho que la edad estimada de la Tierra sea próxima a este periodo de tiempo.[20] La Luna tiene la misma composición que la corteza terrestre pero no contiene un núcleo rico en hierro, como el de la Tierra. Muchos científicos creen que unos 40 millones de años más tarde, un planetoide chocó contra la Tierra, poniendo en órbita material de la corteza terrestre, el cual formó la Luna. Otra hipótesis dice que la Tierra y la Luna comenzaron a juntarse al mismo tiempo, pero la Tierra, con la gravedad mucho más fuertes, atrajo a casi todas las partículas de hierro del área.[21]
Hasta hace poco las rocas más antiguas encontradas en la Tierra eran de hace 3800 millones de años,[20] lo cual hace que lo científicos se inclinen a creer que durante décadas la superficie de la Tierra había sido fundida hasta entonces. En consecuencia, llamaron a esta parte de la historia de la Tierra en el eón Hádico, cuyo nombre significa "infierno".[22] Sin embargo el análisis de circones formados entre 4400 a 4000 millones de años indica que la corteza terrestre se solidificó 100 millones de años después de la formación del planeta y que el planeta rápidamente adquirió los océanos y la atmósfera, que pudieron haber sido capaces de soportar la vida.[23]
La evidencia en la Luna indica que entre hace 4000 a 3800 millones de años atrás sufrió un bombardeo intenso tardío de los escombros que quedaron de la formación del Sistema Solar, y la Tierra debió haber experimentado un bombardeo más intenso debido a su gravedad más fuerte.[22][24] Si bien no hay evidencia directa de las condiciones en la Tierra de entre hace 4000 a 3800 millones de años atrás, no hay ninguna razón para pensar que la Tierra no se vio afectada por este bombardeo intenso tardío.[25] Este evento podría haber despojado a cualquier atmósfera y océanos que existiesen en ese momento; en este caso los gases y el agua de los impactos de cometas pudieron haber contribuido a su reemplazo, aunque también pudo haber contribuido al menos en la mitad la emisión de gases volcánicos en la Tierra.[26]
Evidencia temprana de presencia de la vida en la Tierra
Los primeros organismos fueron identificados en un corto periodo de tiempo y relativamente sin rasgos, sus fósiles parecen pequeñas varillas, que son muy difíciles de distinguir de las estructuras que surgen a través de procesos físicos abióticos. La más antigua evidencia indiscutible de vida en la Tierra, interpretadas como bacterias fosilizadas, datan de hace 3770 millones de años-4280 millones de años.[27] con evidencia geoquímica también parece demostrar la presencia de la vida hace 3800 millones de años.[28] Sin embargo, estos análisis fueron examinados de cerca, y no se encontraron procesos no-biológicos que pudieran producir todos los "signos de vida" de los que se han informado.[29][30] Mientras que esto no pruebe que las estructuras encontradas tengan un origen no biológico, no puede ser tomado como una clara evidencia de la presencia de vida. Marcas geoquímicas en las rocas depositadas hace 3400 millones de años han sido interpretados como evidencia de vida,[31][32] aunque estas declaraciones no han sido completamente examinado por críticos.
Origen de la vida
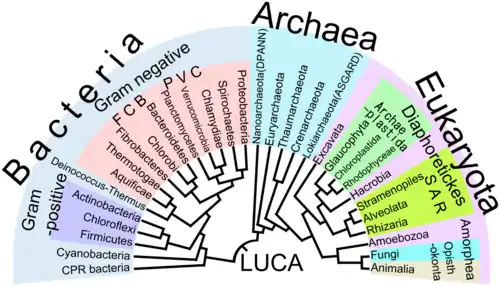
La razón biológica por la que todos los organismos vivos en la Tierra deben compartir el único último antepasado común universal, es porque sería prácticamente imposible que dos o más linajes separados pudieran haber desarrollado de manera independiente los muchos complejos mecanismos bioquímicos comunes a todos los organismos vivos.[34][35] Se ha mencionado anteriormente que las bacterias son los primeros organismos en los que la evidencia fósil está disponible, las células son demasiado complejas para haber surgido directamente de los materiales no vivos.[36] La falta de evidencia geoquímica o fósil de organismos anteriores, o pruebas contundentes de su aparición a partir de protobiontes, ha dejado un amplio campo libre para las hipótesis, que se dividen en dos ideas principales: 1) Que la vida surgió espontáneamente en la Tierra. 2) Que esta fue "sembrada" de otras partes del universo.
Hipótesis de la vida "sembrada" desde otros lugares
La idea de que la vida en la Tierra fue "sembrada" de otras partes del universo se remonta al menos al siglo V a. C.[37] Esto fue propuesto en el siglo XX por el Fisicoquímico Svante Arrhenius,[38] por los astrónomos Fred Hoyle y Chandra Wickramasinghe,[39] y por el biólogo molecular Francis Crick y el químico Leslie Orgel.[40] Hay tres versiones principales de la hipótesis "semilla de otros lugares": 1) En otras partes de nuestro sistema solar a través de choques de fragmentos en el espacio por el impacto de un gran meteorito, en cuyo caso la única fuente creíble es Marte;[41] 2) Por visitantes extraterrestres, posiblemente como resultado de una contaminación interplanetaria accidental por microorganismos que trajeron con ellos,[40] 3) Fuera del sistema solar, pero por medios naturales.[38][41] Los experimentos sugieren que algunos microorganismos pueden sobrevivir al shock de ser catapultados dentro del espacio y también que algunos pueden sobrevivir a la exposición a la radiación durante varios días, pero no hay ninguna prueba de que puedan sobrevivir en el espacio por períodos mucho más largos.[41] Los científicos creen principalmente en dos ideas; sobre la probabilidad de que la vida surgiera de forma independiente en Marte,[42] o en otros planetas en nuestra galaxia.[41]
Hipótesis de la aparición independiente de la vida en la Tierra
La vida en la Tierra está basada en el carbono y el agua. El carbono proporciona un sistema estable para las sustancias químicas complejas y pueden ser fácilmente extraídos del medio ambiente, sobre todo del dióxido de carbono. El único elemento diferente con propiedades químicas similares, es el silicio, este forma estructuras mucho menos estables y, ya que la mayoría de sus compuestos son sólidos, sería más difícil de extraer para los organismos. El agua es un excelente solvente y tiene otras dos propiedades útiles: el hecho de que el hielo flota le permite a los organismos acuáticos sobrevivir debajo del hielo en invierno, y sus moléculas son eléctricamente positivas y negativas, lo que le permite formar una gama más amplia de compuestos de lo que otros solventes pueden tener. Otros buenos solventes, como el amoniaco, sólo son líquidos a temperaturas tan bajas que las reacciones químicas pueden ser demasiado lentas para sustentar la vida y carecen de otras ventajas que posee el agua.[43] Organismos basados en la bioquímica alternativa puede ser de cualquier manera posible en otros planetas.[44]
La investigación sobre cómo la vida pudo haber surgido sin la ayuda de químicos no vivos (protobionte) se centra en tres puntos de partida posibles: autorreplicación, la capacidad de un organismo para producir crías que son muy similares a sí misma; el metabolismo, capacidad para alimentarse y repararse a sí mismo; y membranas plasmáticas, lo que permite que los alimentos entren y que salgan los desechos, pero excluye las sustancias no deseadas.[45] La investigación sobre la abiogénesis todavía tiene un largo camino por recorrer, ya que los enfoques teóricos y empíricos están empezando a entrar en contacto unos con otros.[46][47]
Primer replicación: Mundo del ARN

Incluso los miembros más sencillos de los tres dominios modernos de la vida utilizan el ADN para grabar sus "recetas" junto con un complejo conjunto de moléculas de ARN y proteínas para "leer" las instrucciones y usarlos para el crecimiento de auto-replicación y mantenimiento. Este sistema es demasiado complejo como para haber surgido directamente de los materiales no vivos.[36] El descubrimiento de que algunas moléculas de ARN pueden catalizar su propia replicación y la construcción de proteínas, lleva a la hipótesis de que las primeras formas de vida se basaban enteramente en el ARN.[48] Estas ribozimas pudieron haber formado un mundo de ARN en los que había individuos, pero no especies, como las mutaciones y la transferencia horizontal de genes que han hecho que los descendientes de cada generación sean bastante propensos a tener genomas diferentes de los que sus progenitores empezaron.[49] El ARN más tarde fue sustituido por el ADN, que es más estable y por lo tanto puede construir más genomas, ampliando la gama de capacidades que un solo organismo puede tener.[49][50][51] Los ribozimas siendo uno de los principales componentes de ribosomas, de las células modernas son "fábricas de proteínas".[52]
Aunque pequeñas moléculas auto-replicantes de ARN se han producido artificialmente en laboratorios,[53] se han planteado dudas acerca de dónde la síntesis biológica de ARN es posible naturalmente.[54] Los primeros "ribozimas" pudieron haber sido formados por simples ácidos nucleicos como el ANP, TNA o GNA, que han sido sustituidos más tarde por el ARN.[55][56]
En el 2003 se propuso que el sulfuro de metal poroso precipitado pudo haber ayudado a la síntesis del ARN a unos 100 grados Celsius (212 °F) y a una presión del fondo oceánico cerca de fuentes hidrotermales. En esta hipótesis, las membranas lipídicas serían los últimos componentes importantes de las células en aparecer y hasta entonces las proto-células serían confinadas a los poros.[57]
Primer metabolismo: Mundo de hierro-sulfuro
Una serie de experimentos a partir de 1997 mostró que las primeras etapas en la formación de proteínas a partir de materiales inorgánicos como el monóxido de carbono y sulfuro de hidrógeno pueden lograrse mediante el uso de sulfuro de hierro y sulfuro de níquel como catalizadores. La mayoría de los pasos requieren temperaturas de 100 grados Celsius (212 °F) y presiones moderadas, aunque requiere una etapa de 250 grados Celsius (482 °F) y una presión equivalente de la que se encuentra en el interior de la litosfera, al menos a 7 kilómetros (4,35 mi) de profundidad. Por lo tanto, se sugirió que la síntesis de proteínas auto-sostenible pudo haber ocurrido cerca de las fuentes hidrotermales.[58]
Primeras membranas: Mundo lípido

Se ha sugerido que la doble pared de "burbujas" de los lípidos, como los que forman las membranas externas de las células pudo haber sido un primer paso esencial.[59] Los experimentos que simulan las condiciones de la Tierra primitiva han reportado la formación de lípidos, y que estos pueden ser formados espontáneamente de liposomas, de doble pared de "burbujas", para luego reproducirse a sí mismas. A pesar de que no son intrínsecamente portadores de información de ácidos nucleicos, estarían sujetos a la selección natural para la longevidad y la reproducción. Los ácidos nucleicos como el ARN pudieron haberse formado con más facilidad dentro de los liposomas de lo que les hubiera tomado fuera de estos.[60]
Teoría de la arcilla
El ARN es complejo y existen dudas sobre si se puede producir de una manera no biológica en la naturaleza.[54] Algunas arcillas, sobre todo la montmorillonita, tienen propiedades que las hacen plausibles aceleradoras del surgimiento de un mundo de ARN: estas crecen por medio de la auto-replicación de su patrón cristalino, están sujetos a una selección natural análoga, como las «especies» de la arcilla que crecen más rápido en un ambiente particular que rápidamente se convierte en dominante; y estas pueden catalizar la formación de moléculas de ARN.[61] Aunque esta idea no ha tenido en un consenso científico, todavía tiene partidarios activos.[62]
Medio ambiente e impacto de la evolución de las alfombras microbianas

Las «alfombras» microbianas son múltiples capas, multi-especies de colonias de bacterias y otros organismos que generalmente sólo tienen unos pocos milímetros de grosor, pero todavía contienen una amplia gama de entornos químicos, cada uno de ellos a favor de un conjunto diferente de microorganismos.[63] Hasta cierto punto, cada alfombra forma su propia cadena alimenticia, pues los subproductos de cada grupo de microorganismos generalmente sirven de "alimento" para los grupos adyacentes.[64]
Los estromatolitos son pilares rechonchos construidos como alfombras microbianas que migran lentamente hacia arriba para evitar ser sofocados por los sedimentos depositados en ellos por el agua.[63] Ha habido un intenso debate acerca de la validez de fósiles que supuestamente tienen más de 3000 millones de años,[65] con los críticos argumentando que los llamados estromatolitos podrían haberse formado por procesos no biológicos.[29] Se han reportado estromatolitos en rocas de hace 3500 millones de años en Australia[66] y recientemente microfósiles en Canadá con una antigüedad de entre 3770 millones de años-4280 millones de años.[27]
En las modernas alfombras bajo el agua, la capa superior consiste a menudo de cianobacterias fotosintéticas que crean un ambiente rico en oxígeno, mientras que la capa inferior es libre de oxígeno y, a menudo dominado por el sulfuro de hidrógeno emitido por los organismos que viven allí.[64] Se estima que la aparición de la fotosíntesis oxigénica por las bacterias en las alfombras, aumentó la productividad biológica por un factor de entre 100 y 1000. El agente reductor utilizada por la fotosíntesis oxigénica, es el agua, pues es mucho más abundante que los agentes geológicos producidos por la reducción requerida de la anterior fotosíntesis no oxigénica.[67] A partir de este punto en adelante, la «vida» misma produce mucho más los recursos que necesita que los procesos geoquímicos.[68] El oxígeno, en ciertos organismos, puede ser tóxico, pues éstos no están adaptados a él, así mismo, en otros organismos que sí lo están, aumenta considerablemente su eficiencia metabólica.[69][70] El oxígeno se convirtió en un componente importante de la atmósfera de la Tierra alrededor de hace 2400 millones de años.[71] A pesar de que los eucariotas pueden haber estado presente mucho antes,[72][73] la oxigenación de la atmósfera es un requisito previo para la evolución de las células eucariotas más complejas, de la cual todos los organismos multicelulares están construidos.[74] El límite entre las capas ricas en oxígeno y el oxígeno libre en alfombras microbianas se eleva cuando la fotosíntesis no actúa durante la noche, y luego desciende, al día siguiente. Esto ha creado una presión de selección para los organismos en esta zona intermedia para adquirir la capacidad de tolerar y utilizar el oxígeno, posiblemente a través de la endosimbiosis, donde un organismo vive dentro de otro y ambos se benefician de su asociación.[4]
Las cianobacterias tienen la mayor "caja de herramientas" bioquímica completa de todos los organismos que forman la alfombra. Por lo tanto estos son los organismos más autosuficientes de este sistema, y se adaptan bien al borrar por su cuenta, tanto como alfombras flotantes como el primer fitoplancton, proporcionando la base de la mayoría de las cadenas tróficas marinas.[4]
Diversificación de eucariotas
| Eukaryota |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Las eucariotas pudieron haber estado presente mucho antes de la oxigenación atmosférica,[72] pero las más modernas eucariotas requieren oxígeno, el cual las mitocondrias usan como combustible para la producción de ATP, el suministro de energía interna de todas las células conocidas.[74] En los 1970s se propuso y, después de muchos debates, fue ampliamente aceptado que los eucariotas surgieron como resultado de una secuencia de endosimbiosis entre procariotas. Según el modelo y los análisis moleculares una alfaproteobacteria parásita intracelular invadió una asgardarquea, pero el ataque fue neutralizado, y el atacante se volvió residente y evolucionó en la mitocondria, una de las quimeras, un protozoo tardío más tarde trató de engullir una cianobacteria fotosintética, pero la víctima sobrevivió en el interior del atacante y la nueva combinación se convirtió en el ancestro de las algas y posteriormente las plantas. Después de que cada endosimbiosis comenzara, los socios ya habrían eliminado la duplicación improductiva de las funciones genéticas de reorganización de su genoma, un proceso que a veces implicaba la transferencia de genes entre ellos.[77][78][79] Otra hipótesis propone que las mitocondrias eran originalmente endosimbiontes que metabolizaban hidrógeno o azufre, y más tarde se convirtieron en consumidores de oxígeno.[80] Por otra parte las mitocondrias podrían haber sido parte del equipo original de los eucariotas.[81]
Aparición de los plástidos
Se cree que los plástidos se formaron a partir de cianobacterias endosimbióticas. La simbiosis se desarrolló alrededor de hace 2100-1900 millones de años y ha permitido que los eucariotas lleven a cabo la fotosíntesis oxigénica[82] Tres linajes evolutivos han surgido desde que los plástidos se nombran de manera diferente: los cloroplastos en algas verdes y plantas, rodoplastos en algas rojas y cianelas en glaucofitas.
Organismos multicelulares y reproducción sexual
Evolución de la reproducción sexual
Lo que define la reproducción sexual en eucariotas es la meiosis y la fecundación. En este tipo de reproducción se da mucha recombinación genética, la descendencia recibe el 50% de los genes de cada padre,[83] en contraste con la reproducción asexual, en la que no hay recombinación. Las bacterias también intercambian ADN por conjugación bacteriana, un proceso cuyos beneficios incluyen la resistencia a antibióticos y a otras toxinas, así como la capacidad de utilizar nuevos metabolitos.[84] Sin embargo, la conjugación no es un medio de reproducción, y no se limita a los miembros de la misma especies; hay casos donde las bacterias transfieren ADN a plantas y a animales.[85]
Por otra parte, la transformación bacteriana es claramente una adaptación para la transferencia de ADN entre bacterias de la misma especie. La transformación bacteriana es un proceso complejo que implica los productos de numerosos genes bacterianos y puede considerarse como una forma bacteriana de sexo.[86][87] Este proceso se produce de forma natural en al menos 67 especies procarióticas (en siete filums diferentes).[88] La reproducción sexual en eucariotas puede haber evolucionado a partir de la transformación bacteriana.[89]
Las desventajas de la reproducción sexual son bien conocidas: la reorganización genética durante la recombinación puede alterar combinaciones favorables de genes; y mientras los machos no aumentan directamente el número de crías en su siguiente generación, una población asexual puede rebasar y desplazar a 50 generaciones de una población sexual que es igual en todos los demás aspectos.[83] Sin embargo, la gran mayoría de animales, plantas, hongos y protistas se reproducen sexualmente. Hay fuertes indicios de que la reproducción sexual evolución temprano en la historia de los eucariotas y que los genes que lo controlan han cambiado muy poco desde entonces.[90] ¿Cómo evolucionó y sobrevivió la reproducción sexual es un rompecabezas sin resolver.[91]
La Hipótesis de la Reina Roja sugiere que la reproducción sexual proporciona protección contra los parásitos, porque es más fácil para los parásitos desarrollarse y superar las defensas de clones genéticamente idénticos que en aquellas especies sexuales que presentan defensas dinámicas, y hay una cierta evidencia experimental sobre esto. Por otra parte, contrariamente a las expectativas de la hipótesis de la Reina Roja, Kathryn A. Hanley et al. encontró que la prevalencia, la abundancia y la intensidad de los ácaros fue significativamente mayor en los geckos sexuales que en los asexuales que comparten el mismo hábitat.[92] Además, el biólogo Matthew Parker, después de revisar numerosos estudios genéticos sobre la resistencia a enfermedades de las plantas, no pudo encontrar un solo ejemplo coherente de que los patógenos son el agente selectivo primario responsable de la reproducción sexual en el huésped.[93]
La «hipótesis de la mutación determinista» (HMD) de Alexey Kondrashov asume que cada organismo tiene más de una mutación perjudicial y los efectos combinados de estas mutaciones son más dañinos que la suma de los daños de cada mutación individual. Si es así, la recombinación sexual de genes reducirá el daño que las mutaciones "malas" hacen a la descendencia y al mismo tiempo eliminar algunas de estas mutaciones del acervo genético mediante el aislamiento en las personas que se pierden rápidamente pues tienen un número de mutaciones malas superior a la media. Sin embargo, la evidencia sugiere que los supuestos de la HMD son inestables, debido a que muchas especies tienen en promedio menos de una mutación dañina por individuo y ninguna especie que ha sido investigada muestra evidencia de sinergia entre las mutaciones dañinas.[83]
La naturaleza aleatoria de la recombinación provoca la abundancia relativa de rasgos alternativos que varían de una generación a otra. Esta deriva genética no es suficiente por sí sola para hacer ventajosa a la reproducción sexual, pero una combinación de la deriva genética y la selección natural puede ser suficiente. Cuando una oportunidad produce combinaciones de buenos rasgos, la selección natural le da una gran ventaja a los linajes en que estos rasgos se vincularon genéticamente. Por otro lado, los beneficios de buenos rasgos son neutralizados si aparecen junto con rasgos negativos. La recombinación sexual de buenos rasgos le da oportunidades para vincularse con otros buenos rasgos, y modelos matemáticos sugieren que esto puede ser más que suficiente para compensar las desventajas de la reproducción sexual.[91]
La función adaptativa del sexo hoy sigue siendo unn gran problema pendiente por resolver de la biología. Los modelos de la competencia para explicar la función de adaptación de las relaciones sexuales fueron revisados por John A. Birdsell y Christopher Wills.[96] Las hipótesis discutidas, sobre todo, dependen de los posibles efectos beneficiosos de la variación genética aleatoria producida por la recombinación genética. Una visión alternativa es que el sexo surgió, y se mantiene, como un proceso para la reparación de daños en el ADN, y que la variación genética producida es un subproducto de vez en cuando beneficioso.[89][97]
Pluricelularidad
Las definiciones más simples de pluricelular, como por ejemplo "con múltiples células", pueden incluir a colonias de cianobacterias como Nostoc. Incluso la definición estándar de "tener el mismo genoma, pero diferentes tipos de células" incluye algunos géneros de algas verdes tales como Volvox, que tiene células que se especializan en la reproducción.[98] La pluricelularidad evolucionó independientemente en organismos tan diversos como las esponjas y otros animales, hongos, plantas, algas pardas, cianobacterias, moho mucilaginoso y mixobacterias.[94][99] En resumen, existe una variedad de organismos que muestran la mayor especialización celular y la variedad de tipos de células, aunque esta aproximación a la evolución de la complejidad biológica podría considerarse como "más antropocéntrica".[100]

Las ventajas iniciales de la pluricelularidad incluirían: compartir más eficientemente los nutrientes que son digeridos fuera de la célula,[102] mayor resistencia frente a depredadores, capacidad de resistir corrientes adhiriéndose a una superficie fija, capacidad de trepar para obtener luz para la fotosíntesis,[103] capacidad de crear un ambiente interior protegido del exterior,[100] e incluso dar la oportunidad a un grupo de células de comportarse "inteligentemente" compartiendo información.[101] Estas características podrían haber dado la oportunidad a otros organismos de diversificarse creando entornos más variados que aquellos creados por redes microbianas.[103]
La pluricelularidad con células diferenciadas es beneficiosa para el organismo en su totalidad pero perjudicial desde el punto de vista de las células individuales, ya que la mayoría de ellas pierde la oportunidad de reproducirse. En un organismo pluricelular asexual, aquellas células que retengan la capacidad de reproducción pueden tomar el control y reducir el organismo a una masa de células indiferenciadas. La reproducción sexual elimina dichas células de futuras generaciones y por lo tanto tiende a ser un prerrequisito para una pluricelularidad compleja [103].
Los indicios disponibles indican que los eucariotas evolucionan mucho antes pero permanecen desapercibidos hasta una rápida diversificación hace alrededor de 1000 millones de años. La capacidad de tomar diversas formas es el único aspecto en el que los eucariotas superan a las bacterias y arqueas, y la reproducción sexual permitió a los eucariotas explotar dicha ventaja, produciendo organismos con múltiples células que difieren en forma y función [103].
Indicios fósiles
Los fósiles de la biota francevillense, datados en 2100 millones de años, corresponden probablemente a los organismos pluricelulares más antiguos.[104] Podrían haber tenido células diferenciadas.[105] Otro de los primeros fósiles pluricelulares, Qingshania,[nota 1] datado en 1700 millones de años, consistiría en células virtualmente idénticas. Las algas rojas conocidas como Bangiomorpha, datadas en 1200 millones de años, son los primeros organismos conocidos que poseen con certeza células differenciadas y especializadas, y son también los organismos conocidos más antiguos que se reproducen sexualmente.[103]
Evolución de los animales
Artículos principales: Animal, Explosion cámbrica, y Biota del periodo Ediacárico.
Los animales son organismos pluricelulares y se distinguen de las algas, plantas y hongos por su falta de pared celular. Todos los animales tienen motilidad, aunque sea solo en algunas etapas de su vida. Todos los animales, con la excepción de las esponjas, presentan cuerpos diferenciados con separación entre diferentes tejidos, incluyendo los músculos, que mueven partes del animal al contraerse, y los tejidos nerviosos, que transmiten y procesan señales.
Véase también
Notas
- Nombrado como en el artículo de Nicholas J. Butterfield publicado en Paleobiology, "Bangiomorpha pubescens n. gen., n. sp.: implications for the evolution of sex, multicellularity, and the Mesoproterozoic/Neoproterozoic radiation of eukaryotes" (Summer 2000). Un pez fósil, también de China, también es conocido como Qingshania. El nombre de uno de los dos tendrá que ser cambiado
Referencias
- (en inglés) Steenhuysen, Julie (20 de mayo de 2009). «Study turns back clock on origins of life on Earth». Reuters. Archivado desde el original el 16 de octubre de 2015. Consultado el 27 de septiembre de 2014.
- Futuyma, Douglas J. (2005). Evolution (en inglés). Sunderland, Massachusetts: Sinuer Associates, Inc. ISBN 0-87893-187-2.
- Hawkings, Stephen (1996). «Life in the Universe» (en inglés). Consultado el 9 de abril de 2017.
- Nisbet, E.G., and Fowler, C.M.R. (7 de diciembre de 1999). «Archaean metabolic evolution of microbial mats». Proceedings of the Royal Society: Biology (en inglés) 266 (1436): 2375. PMC 1690475. doi:10.1098/rspb.1999.0934. Consultado el 16 de julio de 2008. «abstracto con relación al contenido completo libre (PDF) ».
- Ariel D. Anbar; Yun Duan, Timothy W. Lyons, Gail L. Arnold, Brian Kendall, Robert A. Creaser, Alan J. Kaufman, Gwyneth W. Gordon, Clinton Scott, Jessica Garvin y Roger Buick (28 de septiembre de 2007). A Whiff of Oxygen Before the Great Oxidation Event? (en inglés) 317 (5846). Science. ISSN 0036-8075. doi:10.1126/science.1140325. Consultado el 30 de agosto de 2011.
- Yin, Leiming (Feb 2020). «Microfossils from the Paleoproterozoic Hutuo Group, Shanxi, North China: Early evidence for eukaryotic metabolism». Precambrian Research 342: 105650. Bibcode:2020PreR..342j5650Y. doi:10.1016/j.precamres.2020.105650.
- Bonner, J.T. (1998) The origins of multicellularity. Integr. Biol. 1, 27–36
- Dr. Paul F. Ciesielski. «Transition of plants to land» (en inglés). University of Florida. Archivado desde el original el 7 de junio de 2011. Consultado el 17 de junio de 2011. «Los fósiles más antiguos revelan la evolución de las plantas no vasculares a mediados y finales de Período Ordovícico (~ 450-440 millones de años) sobre la base de esporas fósiles ».
- Algeo, T.J.; Scheckler, S. E. (1998). «Terrestrial-marine teleconnections in the Devonian: links between the evolution of land plants, weathering processes, and marine anoxic events». Philosophical Transactions of the Royal Society B: Biological Sciences 353 (1365): 113-130. doi:10.1098/rstb.1998.0195.
- «Metazoa: Fossil Record» (en inglés).
- Shu et al.; Luo, H-L.; Conway Morris, S.; Zhang, X-L.; Hu, S-X.; Chen, L.; Han, J.; Zhu, M. et al. (4 de noviembre de 1999). «Lower Cambrian vertebrates from south China». Nature (en inglés) 402 (6757): 42-46. Bibcode:1999Natur.402...42S. doi:10.1038/46965.
- Hoyt, Donald F. (1997). «Synapsid Reptiles» (en inglés). Archivado desde el original el 20 de mayo de 2009.
- Barry, Patrick L. (28 de enero de 2002). «The Great Dying». Science@NASA (en inglés). Science and Technology Directorate, Marshall Space Flight Center, NASA. Archivado desde el original el 16 de febrero de 2012. Consultado el 26 de marzo de 2009.
- Tanner LH, Lucas SG & Chapman MG (2004). «Assessing the record and causes of Late Triassic extinctions» (PDF). Earth-Science Reviews 65 (1-2): 103-139. Bibcode:2004ESRv...65..103T. doi:10.1016/S0012-8252(03)00082-5. Archivado desde el original el 25 de octubre de 2007. Consultado el 22 de octubre de 2007.
- Benton, M.J. (2004). Vertebrate Paleontology (en inglés). Blackwell Publishers. xii-452. ISBN 0-632-05614-2.
- «Amniota - Palaeos» (en inglés). Archivado desde el original el 8 de julio de 2012. Consultado el 18 de junio de 2011.
- Fastovsky DE, Sheehan PM (2005). «The extinction of the dinosaurs in North America». GSA Today (en ingp style="text-indent:-2em">lés) 15 (3): 4-10. doi:10.1130/1052-5173(2005)015<4:TEOTDI>2.0.CO;2. Archivado desde el original el 9 de diciembre de 2011. Consultado el 18 de mayo de 2007.
- «Dinosaur Extinction Spurred Rise of Modern Mammals» (en inglés). News.nationalgeographic.com. Consultado el 8 de marzo de 2009.
- Van Valkenburgh, B. (1999). «Major patterns in the history of carnivorous mammals». Annual Review of Earth and Planetary Sciences (en inglés) 26: 463-493. doi:10.1146/annurev.earth.27.1.463. Archivado desde el original el 17 de febrero de 2021. Consultado el 18 de junio de 2011.
-
Véase:
- Dalrymple, G.B. (1991). The Age of the Earth (en inglés). California: Stanford University Press. ISBN 0-8047-1569-6.
- Newman, W.L. (July de 2007). «Age of the Earth» (en inglés). Publications Services, USGS. Consultado el 29 de agosto de 2008.
- Dalrymple, G.B. (2001). «The age of the Earth in the twentieth century: a problem (mostly) solved». Geological Society, London, Special Publications (en inglés) 190: 205-221. doi:10.1144/GSL.SP.2001.190.01.14. Consultado el 20 de septiembre de 2007.
- Galimov, E.M. and Krivtsov, A.M. (December de 2005). «Origin of the Earth-Moon System». J. Earth Syst. Sci. (en inglés) 114 (6): 593-600. Bibcode:2005JESS..114..593G. doi:10.1007/BF02715942.
- Cohen, B.A., Swindle, T.D. and Kring, D.A. (December de 2000). «Support for the Lunar Cataclysm Hypothesis from Lunar Meteorite Impact Melt Ages». Science (en inglés) 290 (5497): 1754-1756. Bibcode:2000Sci...290.1754C. PMID 11099411. doi:10.1126/science.290.5497.1754. Consultado el 31 de agosto de 2008.
- Véase:
- «Early Earth Likely Had Continents And Was Habitable» (en inglés). University of Colorado. 17 de noviembre de 2005. Archivado desde el original el 14 de octubre de 2008. Consultado el 11 de enero de 2009.
- Cavosie, A.J., Valley, J.W., Wilde, S. A. and the Edinburgh Ion Microprobe Facility (15 de julio de 2005). «Magmatic δ18O in 4400-3900 Ma detrital zircons: A record of the alteration and recycling of crust in the Early Archean». Earth and Planetary Science Letters (en inglés) 235 (3-4): 663-681. Bibcode:2005E&PSL.235..663C. doi:10.1016/j.epsl.2005.04.028.
- Britt, R.R. (24 de julio de 2002). «Evidence for Ancient Bombardment of Earth» (en inglés). Space.com. Archivado desde el original el 4 de octubre de 2002. Consultado el 15 de abril de 2006.
- Valley, J.W., Peck, W.H., King, E.M. and Wilde, S.A. (April de 2002). «A cool early Earth» (PDF). Geology (en inglés) 30 (4): 351-354. doi:10.1130/0091-7613(2002)030<0351:ACEE>2.0.CO;2. Consultado el 13 de septiembre de 2008.
- Dauphas, N., Robert, F. and Marty, B. (December de 2000). «The Late Asteroidal and Cometary Bombardment of Earth as Recorded in Water Deuterium to Protium Ratio». Icarus (en inglés) 148 (2): 508-512. Bibcode:2000Icar..148..508D. doi:10.1006/icar.2000.6489.
- Dodd, Matthew S.; Papineau, Dominic; Grenne, Tor; Slack, John F.; Rittner, Martin; Pirajno, Franco; O’Neil, Jonathan; Little, Crispin T. S. (2 de marzo de 2017). «Evidence for early life in Earth’s oldest hydrothermal vent precipitates». Nature (en inglés) 543 (7643): 60-64. ISSN 0028-0836. doi:10.1038/nature21377. Consultado el 13 de junio de 2017.
- Mojzsis, S.J., Arrhenius, G., McKeegan, K.D., Harrison, T.M., Nutman, A.P. and Friend, C.R.L. (November de 1996). «Evidence for life on Earth before 3,800 million years ago». Nature (en inglés) 384 (6604): 55-59. PMID 8900275. doi:10.1038/384055a0. Consultado el 30 de agosto de 2008.
- Grotzinger, J.P. and Rothman, D.H. (1996). «An abiotic model for stomatolite morphogenesis». Nature (en inglés) 383: 423-425. doi:10.+1038/383423a0.
- Véase:
- Fedo, C.M. and Whitehouse, M.J. (May de 2002). «Metasomatic Origin of Quartz-Pyroxene Rock, Akilia, Greenland, and Implications for Earth's Earliest Life». Science (en inglés) 296 (5572): 1448-1452. PMID 12029129. doi:10.1126/science.1070336. Consultado el 30 de agosto de 2008.
- Lepland, A., van Zuilen, M.A., Arrhenius, G., Whitehouse, M.J. and Fedo, C.M. (January de 2005). «Questioning the evidence for Earth's earliest life — Akilia revisited». Geology (en inglés) 33 (1): 77-79. doi:10.1130/G20890.1. Consultado el 30 de agosto de 2008.
- Brasier, M., McLoughlin, N., Green, O. and Wacey, D. (June de 2006). «A fresh look at the fossil evidence for early Archaean cellular life» (PDF). Philosophical Transactions of the Royal Society: Biology (en inglés) 361 (1470): 887-902. PMC 1578727. PMID 16754605. doi:10.1098/rstb.2006.1835. Consultado el 30 de agosto de 2008.
- Schopf, J. (2006). «Fossil evidence of Archaean life». Philosophical Transactions of the Royal Society of London: B Biological Sciences (en inglés) 361 (1470): 869-85. PMC 1578735. PMID 16754604. doi:10.1098/rstb.2006.1834.
- Ciccarelli, F.D., Doerks, T., von Mering, C., Creevey, C.J., et al (2006). «Toward automatic reconstruction of a highly resolved tree of life». Science (en inglés) 311 (5765): 1283-7. PMID 16513982. doi:10.1126/science.1123061.
- Mason, S.F. (1984). «Origins of biomolecular handedness». Nature (en inglés) 311 (5981): 19-23. PMID 6472461. doi:10.1038/311019a0.
- Orgel, L.E. (October de 1994). «The origin of life on the earth» (PDF). Scientific American (en inglés) 271 (4): 76-83. PMID 7524147. doi:10.1038/scientificamerican1094-76. Consultado el 30 de agosto de 2008. También disponible en esta página web
- Cowen, R. (2000). History of Life (en inglés) (3rd edición). Blackwell Science. p. 6. ISBN 0632044446.
- O'Leary, M.R. (2008). Anaxagoras and the Origin of Panspermia Theory (en inglés). iUniverse, Inc. ISBN 0595495966.
- Arrhenius, S. (1903). «The Propagation of Life in Space». Die Umschau volumen=7. Reprinted in Goldsmith, D., (ed.). The Quest for Extraterrestrial Life (en inglés). University Science Books. ISBN 0198557043.
- Hoyle, F. and Wickramasinghe, C. (1979). «On the Nature of Interstellar Grains». Astrophysics and Space Science (en inglés) 66: 77-90. Bibcode:1979Ap&SS..66...77H. doi:10.1007/BF00648361.
- Crick, F.H.; Orgel, L.E. (1973). «Directed Panspermia». Icarus (en inglés) 19: 341-348. Bibcode:1973Icar...19..341C. doi:10.1016/0019-1035(73)90110-3.
- Warmflash, D. and Weiss, B. (November de 2005). «Did Life Come From Another World?». Scientific American (en inglés): 64-71. Consultado el 2 de septiembre de 2008.
- Ker, Than (August de 2007). «Claim of Martian Life Called 'Bogus'» (en inglés). space.com. Consultado el 2 de septiembre de 2008.
- Bennett, J. O. (2008). «What is life?». Beyond UFOs: The Search for Extraterrestrial Life and Its Astonishing Implications for Our Future (en inglés). Princeton University Press. pp. 82-85. ISBN 0691135495. Consultado el 11 de enero de 2009.
- Schulze-Makuch, D., Irwin, L. N. (April de 2006). «The prospect of alien life in exotic forms on other worlds». Naturwissenschaften (en inglés) 93 (4): 155-72. PMID 16525788. doi:10.1007/s00114-005-0078-6.
- Peretó, J. (2005). «Controversies on the origin of life» (PDF). Int. Microbiol. (en inglés) 8 (1): 23-31. PMID 15906258. Archivado desde el original el 24 de agosto de 2015. Consultado el 7 de octubre de 2007.
- Szathmáry, E. (February de 2005). «Life: In search of the simplest cell». Nature (en inglés) 433 (7025): 469-470. PMID 15690023. doi:10.1038/433469a. Consultado el 1 de septiembre de 2008.
- Luisi, P. L., Ferri, F. and Stano, P. (2006). «Approaches to semi-synthetic minimal cells: a review». Naturwissenschaften (en inglés) 93 (1): 1-13. PMID 16292523. doi:10.1007/s00114-005-0056-z.
- Joyce, G.F. (2002). «The antiquity of RNA-based evolution». Nature (en inglés) 418 (6894): 214-21. PMID 12110897. doi:10.1038/418214a.
- Hoenigsberg, H. (December 2003)). «Evolution without speciation but with selection: LUCA, the Last Universal Common Ancestor in Gilbert’s RNA world». Genetic and Molecular Research (en inglés) 2 (4): 366-375. PMID 15011140. Consultado el 30 de agosto de 2008.(also available as PDF)
- Trevors, J. T. and Abel, D. L. (2004). «Chance and necessity do not explain the origin of life». Cell Biol. Int. (en inglés) 28 (11): 729-39. PMID 15563395. doi:10.1016/j.cellbi.2004.06.006.
- Forterre, P., Benachenhou-Lahfa, N., Confalonieri, F., Duguet, M., Elie, C. and Labedan, B. (1992). «The nature of the last universal ancestor and the root of the tree of life, still open questions». BioSystems (en inglés) 28 (1-3): 15-32. PMID 1337989. doi:10.1016/0303-2647(92)90004-I.
- Cech, T.R. (August de 2000). «The ribosome is a ribozyme». Science (en inglés) 289 (5481): 878-9. PMID 10960319. doi:10.1126/science.289.5481.878. Consultado el 1 de septiembre de 2008.
- Johnston, W. K. et al (2001). «RNA-Catalyzed RNA Polymerization: Accurate and General RNA-Templated Primer Extension». Science (en inglés) 292 (5520): 1319-1325. PMID 11358999. doi:10.1126/science.1060786.
- Véase:
- Levy, M. and Miller, S.L. (July de 1998). «The stability of the RNA bases: implications for the origin of life». Proc. Natl. Acad. Sci. U.S.A. (en inglés) 95 (14): 7933-8. PMC 20907. PMID 9653118. doi:10.1073/pnas.95.14.7933.
- Larralde, R., Robertson, M. P. and Miller, S. L. (August de 1995). «Rates of decomposition of ribose and other sugars: implications for chemical evolution». Proc. Natl. Acad. Sci. U.S.A. (en inglés) 92 (18): 8158-60. PMC 41115. PMID 7667262. doi:10.1073/pnas.92.18.8158.
- Lindahl, T. (April de 1993). «Instability and decay of the primary structure of DNA». Nature 362 (6422): 709-15. PMID 8469282. doi:10.1038/362709a0.
- Orgel, L. (November de 2000). «Origin of life. A simpler nucleic acid». Science (journal) (en inglés) 290 (5495): 1306-7. PMID 11185405. doi:10.1126/science.290.5495.1306.
- Nelson, K.E., Levy, M., and Miller, S.L. (April de 2000). «Peptide nucleic acids rather than RNA may have been the first genetic molecule». Proc. Natl. Acad. Sci. U.S.A. (en inglés) 97 (8): 3868-71. PMC 18108. PMID 10760258. doi:10.1073/pnas.97.8.3868.
- Martin, W. and Russell, M.J. (2003). «On the origins of cells: a hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells». Philosophical Transactions of the Royal Society: Biological (en inglés) 358 (1429): 59-85. PMC 1693102. PMID 12594918. doi:10.1098/rstb.2002.1183. Consultado el 5 de septiembre de 2008.
- Wächtershäuser, G. (August de 2000). «Origin of life. Life as we don't know it». Science (journal) (en inglés) 289 (5483): 1307-8. PMID 10979855. doi:10.1126/science.289.5483.1307.
- Trevors, J.T. and Psenner, R. (2001). «From self-assembly of life to present-day bacteria: a possible role for nanocells». FEMS Microbiol. Rev. (en inglés) 25 (5): 573-82. PMID 11742692. doi:10.1111/j.1574-6976.2001.tb00592.x.
- Segré, D., Ben-Eli, D., Deamer, D. and Lancet, D. (February–April 2001). «The Lipid World» (PDF). Origins of Life and Evolution of Biospheres 2001 (en inglés) 31 (1-2): 119-45. PMID 11296516. doi:10.1023/A:1006746807104. Archivado desde el original el 11 de septiembre de 2008. Consultado el 1 de septiembre de 2008.
- Cairns-Smith, A.G. (1968). «An approach to a blueprint for a primitive organism». En Waddington, C,H., ed. Towards a Theoretical Biology (en inglés) 1. Edinburgh University Press. pp. 57-66.
- Ferris, J.P. (junio de 1999). «Prebiotic Synthesis on Minerals: Bridging the Prebiotic and RNA Worlds». Biological Bulletin. Evolution: A Molecular Point of View (en inglés) (Biological Bulletin, Vol. 196, No. 3) 196 (3): 311-314. JSTOR 1542957. PMID 10390828. doi:10.2307/1542957. Consultado el 1 de septiembre de 2008.
- Krumbein, W.E., Brehm, U., Gerdes, G., Gorbushina, A.A., Levit, G. and Palinska, K.A. (2003). «Biofilm, Biodictyon, Biomat Microbialites, Oolites, Stromatolites, Geophysiology, Global Mechanism, Parahistology» (PDF). En Krumbein, W.E., Paterson, D.M., and Zavarzin, G.A., ed. Fossil and Recent Biofilms: A Natural History of Life on Earth (en inglés). Kluwer Academic. p. 1–28. ISBN 1402015976. Archivado desde el original el 6 de enero de 2007. Consultado el 9 de julio de 2008.
- Risatti, J. B., Capman, W. C. and Stahl, D. A. (11 de octubre de 1994). «Community structure of a microbial mat: the phylogenetic dimension» (PDF). Proceedings of the National Academy of Sciences (en inglés) 91 (21): 10173-10177. Bibcode:1994PNAS...9110173R. PMC 44980. PMID 7937858. doi:10.1073/pnas.91.21.10173. Consultado el 9 de julio de 2008.
- (the editor) (June 2006)). «Editor's Summary: Biodiversity rocks». Nature (en inglés) 441 (7094). Consultado el 10 de enero de 2009.
- Allwood, A. C., Walter, M. R., Kamber, B. S., Marshall, C. P. and Burch, I. W. (June 2006)). «Stromatolite reef from the Early Archaean era of Australia». Nature (en inglés) 441 (7094): 714–718. Bibcode:2006Natur.441..714A. PMID 16760969. doi:10.1038/nature04764. Consultado el 31 de agosto de 2008.
- Blankenship, R.E.; Bebout, BM; Des Marais, DJ (1 de enero de 2001). «Molecular evidence for the evolution of photosynthesis». Trends in Plant Science (en inglés) 6 (1): 4–6. PMID 11460161. doi:10.1038/35085554. Consultado el 14 de julio de 2008.
- Hoehler, T.M., Bebout, B.M. and Des Marais, D.J. (19 de julio de 2001). «The role of microbial mats in the production of reduced gases on the early Earth». Nature (en inglés) 412 (6844): 324–327. PMID 11460161. doi:10.1038/35085554. Consultado el 14 de julio de 2008.
- Abele, D. (7 de noviembre de 2002). «Toxic oxygen: The radical life-giver». Nature (en inglés) 420 (27): 27. PMID 12422197. doi:10.1038/420027a. Consultado el 14 de julio de 2008.
- «Introduction to Aerobic Respiration» (en inglés). University of California, Davis. Archivado desde el original el 29 de octubre de 2007. Consultado el 14 de julio de 2008.
- Goldblatt, C., Lenton, T.M. and Watson, A.J. (2006). «The Great Oxidation at ~2.4 Ga as a bistability in atmospheric oxygen due to UV shielding by ozone» (PDF). Geophysical Research Abstracts (en inglés) 8 (00770). Consultado el 1 de septiembre de 2008.
- Glansdorff, N., Xu, Y. and Labedan, B. (2008). «The Last Universal Common Ancestor: emergence, constitution and genetic legacy of an elusive forerunner». Biology Direct (en inglés) 3 (29): 29. PMC 2478661. PMID 18613974. doi:10.1186/1745-6150-3-29.
- Brocks, J. J., Logan, G. A., Buick, R. and Summons, R. E. (1999). «Archaean molecular fossils and the rise of eukaryotes». Science (en inglés) 285 (5430): 1033–1036. PMID 10446042. doi:10.1126/science.285.5430.1033. Consultado el 2 de septiembre de 2008.
- Hedges, S. B., Blair, J. E., Venturi, M. L. and Shoe, J. L (January de 2004). «A molecular timescale of eukaryote evolution and the rise of complex multicellular life». BMC Evolutionary Biology (en inglés) 4 (2): 2. PMC 341452. PMID 15005799. doi:10.1186/1471-2148-4-2. Consultado el 14 de julio de 2008.
- Brown, M. W., Heiss, A. A., Kamikawa, R., Inagaki, Y., Yabuki, A., Tice, A. K., ... & Roger, A. J. (2018). Phylogenomics places orphan protistan lineages in a novel eukaryotic super-group. Genome biology and evolution, 10(2), 427-433.
- Romain Derelle et al 2015, Bacterial proteins pinpoint a single eukaryotic root. Proceedings of the National Academy of Sciences 112(7) · January 2015 with 248 Reads DOI: 10.1073/pnas.1420657112
- Margulis, L. (1981). Symbiosis in cell evolution (en inglés). San Francisco: W.H. Freeman. ISBN 0-7167-1256-3.
- Vellai, T. and Vida, G. (1999). «The origin of eukaryotes: the difference between prokaryotic and eukaryotic cells». Proceedings of the Royal Society: Biology (en inglés) 266 (1428): 1571-1577. PMC 1690172. PMID 10467746. doi:10.1098/rspb.1999.0817.
- Selosse, M-A., Abert, B., and Godelle, B. (2001). «Reducing the genome size of organelles favours gene transfer to the nucleus». Trends in ecology & evolution (en inglés) 16 (3): 135-141. doi:10.1016/S0169-5347(00)02084-X. Consultado el 2 de septiembre de 2008.
- Pisani, D., Cotton, J.A. and McInerney, J.O. (2007). «Supertrees disentangle the chimerical origin of eukaryotic genomes». Mol Biol Evol. (en inglés) 24 (8): 1752-60. PMID 17504772. doi:10.1093/molbev/msm095.
- Gray, M.W., Burger, G., and Lang, B.F. (1999). «Mitochondrial evolution». Science (en inglés) 283 (5407): 1476-1481. Bibcode:1999Sci...283.1476G. PMID 10066161. doi:10.1126/science.283.5407.1476. Consultado el 2 de septiembre de 2008.
- Jürgen F. H. Strassert, Iker Irisarri, Tom A. Williams & Fabien Burki (2021). A molecular timescale for eukaryote evolution with implications for the origin of red algal-derived plastids.
- Neiman, Maurine; Jokela, Jukka (2010). «Sex: Advantage». Encyclopedia of Life Sciences. Hoboken, NJ: John Wiley & Sons. ISBN 0-470-01617-5. doi:10.1002/9780470015902.a0001716.pub2. Consultado el 21 de enero de 2015.
- Holmes y Jobling, 1996
- Christie, Peter J. (April 2001). «Type IV secretion: intercellular transfer of macromolecules by systems ancestrally related to conjugation machines». Molecular Microbiology (Hoboken, NJ: Wiley-Blackwell) 40 (2): 294-305. ISSN 0950-382X. PMID 11309113. doi:10.1046/j.1365-2958.2001.02302.x.
- Michod, Richard E.; Bernstein, Harris; Nedelcu, Aurora M. (May 2008). «Adaptive value of sex in microbial pathogens» (PDF). Infection, Genetics and Evolution (Amsterdam, the Netherlands: Elsevier) 8 (3): 267-285. ISSN 1567-1348. PMID 18295550. doi:10.1016/j.meegid.2008.01.002.
- Bernstein, Harris; Bernstein, Carol (July 2010). «Evolutionary Origin of Recombination during Meiosis». BioScience (Oxford: Oxford University Press on behalf of the American Institute of Biological Sciences) 60 (7): 498-505. ISSN 0006-3568. doi:10.1525/bio.2010.60.7.5.
- Johnsborg, Ola; Eldholm, Vegard; Håvarstein, Leiv Sigve (December 2007). «Natural genetic transformation: prevalence, mechanisms and function». Research in Microbiology (Amsterdam, the Netherlands: Elsevier for the Pasteur Institute) 158 (10): 767-778. ISSN 0923-2508. PMID 17997281. doi:10.1016/j.resmic.2007.09.004.
- Bernstein, Bernstein y Michod, 2012, pp. 1-50
- Ramesh, Marilee A.; Malik, Shehre-Banoo; Logsdon, John M., Jr. (26 de enero de 2005). «A Phylogenomic Inventory of Meiotic Genes: Evidence for Sex in Giardia and an Early Eukaryotic Origin of Meiosis» (PDF). Current Biology (Cambridge, MA: Cell Press) 15 (2): 185-191. ISSN 0960-9822. PMID 15668177. doi:10.1016/j.cub.2005.01.003. Archivado desde el original el 26 de febrero de 2009. Consultado el 22 de diciembre de 2008.
- Otto, Sarah P.; Gerstein, Aleeza C. (August 2006). «Why have sex? The population genetics of sex and recombination». Biochemical Society Transactions (London: Portland Press on behalf of the Biochemical Society) 34 (Pt 4): 519-522. ISSN 0300-5127. PMID 16856849. doi:10.1042/BST0340519. Consultado el 22 de diciembre de 2008.
- Hanley, Kathryn A.; Fisher, Robert N.; Case, Ted J. (June 1995). «Lower Mite Infestations in an Asexual Gecko Compared With Its Sexual Ancestors». Evolution (Hoboken, NJ: John Wiley & Sons on behalf of the Society for the Study of Evolution) 49 (3): 418-426. ISSN 0014-3820. doi:10.2307/2410266.
- Parker, Matthew A. (September 1994). «Pathogens and sex in plants». Evolutionary Ecology (Dordrecht, the Netherlands: Kluwer Academic Publishers) 8 (5): 560-584. ISSN 0269-7653. doi:10.1007/bf01238258.
- Fedonkin, Mikhail A. (31 de marzo de 2003). «The origin of the Metazoa in the light of the Proterozoic fossil record» (PDF). Paleontological Research (Tokyo: Palaeontological Society of Japan) 7 (1): 9-41. ISSN 1880-0068. doi:10.2517/prpsj.7.9. Archivado desde el original el 11 de septiembre de 2008. Consultado el 2 de septiembre de 2008.
- Birdsell y Wills, 2003, pp. 27-137
- Bernstein, Hopf y Michod, 1987, pp. 323–370
- Bell, Graham; Mooers, Arne O. (1997). «Size and complexity among multicellular organisms» (PDF). Biological Journal of the Linnean Society (Hoboken, NJ: John Wiley & Sons on behalf of the Linnean Society of London) 60 (3): 345-363. ISSN 0024-4074. doi:10.1111/j.1095-8312.1997.tb01500.x. Consultado el 2 de febrero de 2015.
- Kaiser, Dale (December 2001). «Building a multicellular organism». Annual Review of Genetics (Palo Alto, CA: Annual Reviews) 35: 103-123. ISSN 0066-4197. PMID 11700279. doi:10.1146/annurev.genet.35.102401.090145.
- Bonner, John Tyler (1998). «The origins of multicellularity». Integrative Biology (London: Royal Society of Chemistry) 1 (1): 27-36. ISSN 1757-9694. doi:10.1002/(SICI)1520-6602(1998)1:1<27::AID-INBI4>3.0.CO;2-6. Consultado el 3 de septiembre de 2008.
- Nakagaki, Toshiyuki; Yamada, Hiroyasu; Tóth, Ágota (28 de septiembre de 2000). «Intelligence: Maze-solving by an amoeboid organism». Nature (London: Nature Publishing Group) 407 (6803): 470. ISSN 0028-0836. PMID 11028990. doi:10.1038/35035159. Consultado el 3 de septiembre de 2008.
- Koschwanez, John H.; Foster, Kevin R.; Murray, Andrew W. (9 de agosto de 2011). «Sucrose Utilization in Budding Yeast as a Model for the Origin of Undifferentiated Multicellularity». PLOS Biology (San Francisco, CA: Public Library of Science) 9 (8): e1001122. ISSN 1545-7885. doi:10.1371/journal.pbio.1001122. Consultado el 2 de febrero de 2015.
- Butterfield, Nicholas J (2000). «Bangiomorpha pubescens n. gen., n. sp.: implications for the evolution of sex, multicellularity, and the Mesoproterozoic/Neoproterozoic radiation of eukaryotes». Paleobiology (Paleontological Society) 26 (3): 386-404. ISSN 0094-8373. doi:10.1666/0094-8373(2000)026<0386:BPNGNS>2.0.CO;2.
- El Albani, Abderrazak; Bengtson, Stefan; Canfield, Donald E. et al. (1 de julio de 2010). «Large colonial organisms with coordinated growth in oxygenated environments 2.1 Gyr ago». Nature (London: Nature Publishing Group) 466 (7302): 100-104. Bibcode:2010Natur.466..100A. ISSN 0028-0836. PMID 20596019. doi:10.1038/nature09166.
- Dickey, Gwyneth (31 de julio de 2010). «Evidence for earlier multicellular life» (PNG). Science News (Washington, D.C.: Society for Science & the Public) 178 (3): 17. ISSN 0036-8423. doi:10.1002/scin.5591780322. Consultado el 2 de febrero de 2015.
- «Eukaryotic organisms in Proterozoic oceans». Philosophical Transactions of the Royal Society of London, Part B (en inglés) 361 (1470): 1023-38. 2006. PMC 1578724. PMID 16754612. doi:10.1098/rstb.2006.1843.
- Rasmussen, B., Fletcher, I.R., Brocks, J.R. and Kilburn, M.R. (October de 2008). «Reassessing the first appearance of eukaryotes and cyanobacteria». Nature (en inglés) 455 (7216): 1101-1104. Bibcode:2008Natur.455.1101R. PMID 18948954. doi:10.1038/nature07381.
- Han, T.M. and Runnegar, B. (July de 1992). «Megascopic eukaryotic algae from the 2.1-billion-year-old negaunee iron-formation, Michigan». Science (en inglés) 257 (5067): 232-235. Bibcode:1992Sci...257..232H. PMID 1631544. doi:10.1126/science.1631544. Consultado el 2 de septiembre de 2008.
- Javaux, E. J., Knoll, A. H. and Walter, M. R. (September de 2004). «TEM evidence for eukaryotic diversity in mid-Proterozoic oceans». Geobiology (en inglés) 2 (3): 121-132. doi:10.1111/j.1472-4677.2004.00027.x. Consultado el 2 de septiembre de 2008.
- Butterfield, N. J. (2005). «Probable Proterozoic fungi». Paleobiology (en inglés) 31 (1): 165-182. ISSN 0094-8373. doi:10.1666/0094-8373(2005)031<0165:PPF>2.0.CO;2. Consultado el 2 de septiembre de 2008.
| Control de autoridades |
|
|---|
 Datos: Q1098275
Datos: Q1098275 Multimedia: Prehistoric life / Q1098275
Multimedia: Prehistoric life / Q1098275