Cerf élaphe
Le cerf élaphe (Cervus elaphus) est un grand cervidé des forêts tempérées d'Europe, d'Afrique du Nord et d'Asie occidentale et centrale. Son nom est un pléonasme, car « élaphe » signifie déjà « cerf » en grec.
Ce cerf est l'un des représentants les plus connus de cette famille de mammifères. L'espèce comprend plusieurs sous-espèces, dont le « cerf de Bactriane », le « cerf du Turkestan », le « cerf rouge du Turkestan » ou encore le « cerf élaphe du Turkestan »[1].
La femelle est la « biche », le petit est le faon. Ensuite, de 6 mois à 1 an, on l'appelle « hère » s'il s'agit d'un mâle, la femelle conserve le nom de « faon » jusqu'à un an. De 1 an à 2 ans, le jeune mâle est appelé « daguet » avec deux grands bois secs et la femelle « bichette ». Les biches ne portent pas de bois.
Cervus elaphus
Pour les articles homonymes, voir Cerf (homonymie).
Ne pas confondre avec le genre Elaphe, des serpents de la famille des colubridés.

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Classe | Mammalia |
| Ordre | Artiodactyla |
| Sous-ordre | Ruminantia |
| Famille | Cervidae |
| Sous-famille | Cervinae |
| Genre | Cervus |

LC : Préoccupation mineure
Statut CITES
Statut CITES
Statut CITES
Répartition géographique
_reconstructed_and_recent.png.webp)
Origine, histoire, nomenclature et systématique
.jpg.webp)
.jpg.webp)

Les cerfs sont de grands herbivores ruminants présents dans l'hémisphère nord. Originellement adaptés aux milieux ouverts enherbés ou à des zones de type savanes tempérées (broutage de feuilles et bourgeons à différentes hauteurs), ils ont formé des populations et sous-populations qui ont évolué, sous la contrainte des dernières glaciations, et plus rapidement ensuite sous la contrainte de la chasse par l'homme et de l’anthropisation des paysages[3]. À la différence du cerf megaloceros, ils ont, sur une grande partie de leur aire potentielle de répartition survécu aux chasseurs du paléolithique et du néolithique en se réfugiant dans les zones de forêt dense. Les grands cervidés doivent cependant affronter localement des phénomènes d'insularisation écologique notamment dû à la fragmentation des forêts où ils se sont réfugiés, ou au fait que certaines populations sont issues d'un petit nombre d'individus réintroduits (risques liés à la consanguinité). Des sous-populations ont pu évoluer de manière assez divergente, notamment dans le contexte d'insularisation écologique des îles méditerranéennes au Pléistocène, avec des variations de taille, mais aussi fonctionnelles (par exemple de l'appareil masticateur ou des os du pied, qui sont probablement des adaptations aux modifications des niches écologiques occupées[4]). Le « mégacérin de la Sardaigne » et le « cerf de Sicile » devaient être des herbivores agiles et rapides. Le mégacérin de Sicile, semble quant à lui avoir été plus adapté à des milieux plus fermés et boisés[4]. En Crète, les petits mégacérins, même peu agiles, devaient être adaptés à des terrains difficiles et accidentés, sans qu'on sache exactement quelle pouvait être leur alimentation[4].
Le cerf élaphe comme toute la grande et petite faune des zones tempérées a récemment (aux échelles géologiques du terme) dû s'adapter à trois glaciations. De premiers échantillons d'ADN bien conservé ont été trouvés sur des os de trois cerfs (Cervus elaphus) datés de la fin du Pléistocène, trouvés dans la grotte Emine-Bair-Khosar (EBK) à la lisière nord du plateau inférieur du Massif montagneux de Tchatyr-Dah en Crimée[5]. En les comparant à l'arbre phylogéographique connu basé sur l'analyse de séquences d'ADN mitochondrial de cerfs vivant aujourd'hui dans l'hémisphère Nord, les chercheurs ont pu confirmer que la péninsule de Crimée semble avoir servi de refuge à une grande part des cerfs élaphes du nord-Est de l'Europe, et qu'elle a ensuite (Pléistocène supérieur) joué un « rôle majeur » dans la recolonisation et la dispersion des espèces tempérées pour tout le continent eurasien[5].
Pour éclairer la phylogénie, la phylogéographie du cerf élaphe, une étude publiée en 2004 a comparé la séquence d'ADN mitochondrial du cytochrome b d'échantillons prélevés parmi cinquante et une populations de cerfs réparties dans le monde, mais essentiellement en Asie et Europe[6].
Plusieurs méthodes ont été combinées (dont le maximum de parcimonie, le maximum de vraisemblance, et l'analyse cladistique imbriquée), qui ont conduit les chercheurs à conclure que les populations de cerf élaphe actuellement présentes seraient originaires de la région située entre le Kirghizistan et l'Inde du Nord, avec deux groupes génétiquement bien distincts, l'un vivant à l'ouest (composé de quatre sous-groupes) et un autre vivant à l'est (composée de trois sous-groupes)[6].
Ces données ADNmt ne confirment pas la classification traditionnelle considérant que tous les cerfs élaphes constituent une seule espèce, ni sa division en nombreuses sous-espèces[6].
Systématique
Dans la littérature antérieure aux nouvelles données génomiques, l'espèce Cervus elaphus compte plusieurs sous-espèces :
- le cerf de Barbarie (Cervus elaphus barbarus), qui n'existe plus que dans une aire très limitée de l'Atlas ;
- le cerf élaphe de Corse et Sardaigne (Cervus elaphus corsicanus) ;
- le maral (Cervus elaphus maral) d'Europe Centrale et de Sibérie ;
- le cerf de Bactriane ou cerf de Boukhara d'Asie centrale (Cervus elaphus yarkandensis, anciennement Cervus elaphus bactrianus) ;
- le hangul (Cervus elaphus hanglu) du Cachemire ;
- le cerf de Roosevelt (Cervus elaphus roosevelti) d'Amérique du Nord et de Sibérie ;
- le cerf blanc du Danemark.
Ces espèces peuvent comprendre plusieurs génotypes ayant permis des adaptations aux caractéristiques biogéographiques et écologiques des zones où elles ont vécu, et leur classification pourrait devoir être revue.
Cette classification est actuellement remise en cause par certains taxonomistes. Après analyse génétique[6], le wapiti (anciennement Cervus elaphus wapiti) est considéré comme une espèce à part entière sous le nom de Cervus canadensis. Le nombre de sous-espèces tendrait à se réduire pour n'en conserver que deux.[réf. nécessaire]
Description et caractéristiques

Données biométriques
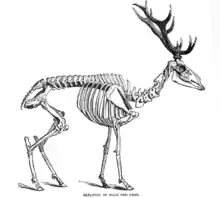


Les cerfs élaphe contemporains sont élancés, ils atteignent une longueur totale de 1,6 à 2,6 mètres, pour 1,10 à 1,50 mètre de hauteur au garrot et un poids de 67 à 300 kilogrammes, variant selon la sous-espèce. Le poids des animaux est variable selon l'âge, le sexe et la région. Le poids des cerfs augmente de l'ouest vers l'est de l'Europe : de 100 kg en Écosse à 250 kg en Europe de l'Est. Le poids des animaux se stabilise vers l'âge de 3–4 ans chez la femelle (biche) et 7 ans chez le mâle.
En France, les cerfs mâles pèsent entre 120 et 250 kg (150 kg en moyenne) pour une taille de 130 à 150 cm (1,40 mètre en moyenne) au garrot et environ 1,70 à 1,80 m à la hauteur de la tête, les biches, pèsent entre 67 et 100 kg (80 kg en moyenne) pour une taille qui varie selon les individus de 1,10 à 1,30 mètre (1,20 mètre en moyenne) au garrot et 1,50 m environ à la hauteur de la tête.
Ce poids varie selon la qualité du milieu de vie (abondance de la nourriture) mais aussi selon la saison. Un hiver rude peut entraîner une perte de poids de 10 à 15 % chez les deux sexes, le mâle peut perdre jusqu'à 20 % de son poids pendant le brame et la femelle jusqu'à 15 % de son poids pendant la période d'allaitement. Les activités physiques également, notamment la course et les bonds, montrent qu'en l'absence de prédateurs, les cerfs se déplacent moins et peuvent prendre plus de poids.
La queue mesure de 10 à 27 centimètres de long. Les sous-espèces nord-américaines et du nord-est de l'Asie sont généralement plus grandes que les européennes. Les mâles sont également toujours plus massifs que les femelles. Le cerf élaphe est élancé, mais fortement constitué, avec un poitrail massif, et un cou assez élancé. Les yeux sont de taille moyenne, les oreilles effilées aussi longues que la moitié de la tête et des pattes très fines adaptées à la course rapide et aux bonds. Les pattes sont constituées de quatre doigts (les doigts 2, 3, 4, 5, le « pouce » étant atrophié), les doigts 2 et 5, plus petits que les deux autres et ramenés vers les côtés de la patte, ne marquent normalement pas le sol lors des déplacements, excepté lors de déplacements dans la neige ou la boue.
On trouvait en Gaule au moins jusqu'à l'époque gallo-romaine des cerfs de très grande taille, comme en témoigne par exemple le squelette entier d'un cerf retrouvé par les archéologues MM. Métayer et Gardin en 1856 dans un puits funéraire gallo-romain. Les mensurations du crâne de ce cerf sont de 56 cm de long et 25 cm de large (entre les deux arcades sourcilières) celles des omoplates : 37 cm de long et 23 cm de large. De nombreux squelettes de loups et d'autres animaux contemporains de ce cerf ont été trouvés dans ce même puits, avec quelques têtes de chiots.
Pelage


La coloration du pelage varie fortement selon les saisons, l'âge et le sexe : d'une teinte brun-roux en été et gris-brun en hiver ; le mâle a généralement un pelage plus sombre que la femelle.
La mue intervient deux fois par an, aux printemps (avril-mai), alors les cerfs perdent leur pelage épais de l'hiver pour laisser apparaître une nouvelle peau plus fine et plus claire, puis à l'automne (octobre) apparaît un pelage plus épais et plus foncé pour supporter les froids d'hiver.
Une tache jaune clair, appelée « cimier », orne la croupe des deux sexes.
Le faon a, jusqu'à l'âge de quatre mois environ, un pelage brun clair tacheté de blanc que l'on appelle « livrée ».
Exceptionnellement, des cerfs ont un pelage blanc, tels qu'en captivité au parc de Dyrehaven (Danemark), et au moulin de Poyaller (dans les Landes, en France)[7] ou à l'élevage de la belle charme en Haute-Marne (Champagne-Ardenne).
Bois


À partir de neuf mois, des bois commencent à pousser sur la tête du mâle sous forme de pivots. À un an, ils sont visibles, et le mâle est alors appelé « daguet ». Les bois vont alors tomber chaque année (fin de l'hiver chez les cerfs âgés, début du printemps chez les jeunes) pour repousser au cours de l'été. Une enveloppe nourricière, duveteuse et irriguée de sang, assure leur croissance à la manière d'un placenta pour un fœtus. Fin juillet, le velours tombe. Paradoxe de la nature, les bois ne sont opérationnels que lorsqu'ils sont morts. Pour les dépouiller totalement puis les aiguiser en vue des prochains combats, le cerf va « frayer » en les frottant de façon répétitive aux arbres. Il mange les lambeaux de peau qui pendent aux ramures.
Les dimensions et la forme des bois varient individuellement, mais aussi selon l'âge et la sous-espèce[8]. Contrairement à une croyance répandue, le nombre de cors n'a pas de rapport direct avec l'âge[9]. Le premier cor se nomme andouiller de massacre, le second surandouiller.
Le bois de cerf râpé ou calciné était employé jusqu'à la fin du XIXe siècle en tant qu'antidiarrhéique, les principes actifs étant la gélatine et le phosphate de calcium[réf. souhaitée].
Répartition
Le groupe des « cerfs élaphes » est répandu dans l’hémisphère nord en trois grandes populations séparées, aujourd'hui classées en deux espèces différentes depuis 2004. L'espèce Cervus elaphus, le cerf élaphe proprement dit, est désormais réduite à la population du Paléarctique occidental seulement. Il est présent dans l'essentiel de l'Europe, le nord-ouest de l'Afrique, dans le nord du Moyen-Orient et en Asie centrale jusqu'aux contreforts sud-est du Tian Shan à l'extrême ouest de la Chine. Tandis que Cervus canadensis, le wapiti, rassemble les populations du Paléarctique oriental (Asie tempérée, jusqu'à l'Altaï à l'ouest) et du Néarctique (Amérique du Nord).
Il a été introduit et est élevé pour sa viande dans certains pays, comme en Nouvelle-Zélande où il est aujourd'hui invasif.
En France, il est aujourd'hui commun ou redevenu commun dans de nombreuses régions. Parmi les nombreux massifs à cerf, citons :
- la Petite Pierre (Bas-Rhin), site d'étude de l'Office national de la chasse et de la faune sauvage ;
- le massif de Châteauvillain- Arc-en-Barrois (Haute-Marne) ;
- Chambord (Loir-et-Cher) (réserve de chasse) ;
- la forêt de Compiègne (Oise) ;
- la forêt de Tronçais (Allier).
Le cerf a parfois été trop chassé ce qui a conduit à sa disparition d'une grande partie de la France, et les populations menacées de certains massifs ont dû être reconstituées par réintroduction de l'espèce (dans la forêt de Paimpont en Bretagne ou de Mormal dans le nord par exemple).
En France, à la suite de la disparition de ses prédateurs naturels et depuis l'instauration des plans de chasse dans les années 1960, la population est en augmentation continue. Ainsi, la population serait passée de 40 000 en 1983 à 100 000 en 1994, 160 000 en 2010 voire plus de 180 000 en 2020. Localement, d'importants dégâts sont constatés sur les jeunes arbres. Toutefois, la capacité maximale des cerfs élaphes en France serait de 700 000 individus.[réf. nécessaire]
Habitat



Il vit surtout dans les grands massifs de la forêt tempérée européenne ainsi que dans les forêts et maquis méditerranéens. Mais dans ces massifs forestiers il affectionne surtout les zones de bois clairs, les parcelles coupées et en régénération, les trouées et clairières avec prairies ou landes et les larges chemins d'exploitation peu fréquentés. Le soir il peut sortir dans les champs et les prairies des lisières s'il y trouve de la tranquillité. Il utilise la forêt dense pour se cacher, notamment la journée. Il est présent aussi dans les forêts de montagne jusqu'à plus de 2 000 m et il fréquente les alpages.
Ses caractéristiques morphologiques et biologiques (animal taillé pour la course, vue et odorat très développés, la taille de ses bois), son régime alimentaire (essentiellement des plantes de lumière qu'il broute) et son comportement (vie en troupeaux pouvant être importants, territorialité en période de rut), indiquent qu'il s'agit plutôt d'un animal de milieu semi-ouvert à ouvert. Mais il s'est réfugié au sein des grands massifs forestiers car il a été chassé des terres agricoles et des prairies pastorales par l'homme depuis des temps très anciens. Il a vécu par exemple en grands troupeaux dans les steppes d'Europe de l'Est, en zones dépourvues de tout boisement, mais il en a aujourd'hui disparu car il était plus facile à trouver et à chasser dans cet habitat. En Écosse, il vit encore actuellement en forte densité dans de vastes landes ouvertes dépourvues de forêt. Dans les Alpes, il séjourne dans les alpages où il peut aller jusqu'à la limite de la végétation à plus 3 000 m d'altitude[10].
Son territoire vital est estimé en France à entre 1 000 et 5 000 ha (soit 10–50 km2) ; 3 000 ha (30 km2) en moyenne, ce chiffre variant considérablement selon la productivité du milieu. Ces animaux se déplacent parfois sur de longues distances (plusieurs dizaines de kilomètres). La fragmentation forestière peut donc leur nuire, mais elle semble pouvoir être compensée par des écoducs judicieusement placés (au-dessus d'autoroutes ou de TGV par exemple).
En Grèce, le cerf élaphe est en danger critique d'extinction[11].
Mode de vie et comportement
Mœurs et traits de vie
Le cerf est une espèce crépusculaire et nocturne (ou qui l'est devenue en s'adaptant à la pression de chasse, comme elle est probablement devenue forestière tout en étant plus adaptée aux milieux semi-ouverts ou ouverts).
L'espèce est grégaire. Les femelles (biches) se regroupent en hardes dont le domaine vital est de 2 000 à 5 000 ha[12]. Les mâles vivent avec elles durant la saison des amours (le rut). Un seul mâle peut être accompagné d'une harde qui peut compter jusqu'à soixante biches. La biche est la seule à prendre soin de son faon. Elle ne le quitte que pour aller brouter. La gestation est d'environ huit mois[13].
Composition sociale
Les groupes ou hardes sont composées de mâles, de femelles ou mixtes. La cellule de base est « matriarcale » (avec un groupe triangulaire composé d'une biche suivie du jeune de l’année et du jeune de l’année précédente).
Le cerf marque son territoire, par le brame du mâle, les excréments, l'abroutissement (plantules mangées, extrémités des rameaux latéraux ou de la pousse terminale des végétaux ligneux rongés), écorçage par les incisives inférieures dans un but alimentaire, peut être « médical » mais aussi comportemental) et frottis du cerf mâle (en période de reproduction et de repousse des bois)[12].
Le rut et la reproduction


Mâles et femelles vivent séparément de décembre à août et se retrouvent pour une période de fécondation qui s'étend en Europe tempérée du au . Le système reproducteur des mâles et des jeunes biches nées de l'année précédente est en état de quiescence jusqu'au moment du rut où sous l'effet de certaines hormones, le sperme du cerf mâle devient plus abondant et change de composition[15], ce qui a une importance pour les banques de sperme qui souhaitent conserver des spermatozoïdes d'espèces menacées de cerfs, ou pour l'insémination artificielle de biches d'élevage[16].
Le rut, marqué par le cri rauque et retentissant du mâle (le brame) intervient à la fin de l'été ou au début de l'automne, et dure environ un mois, mais on peut encore entendre bramer des cerfs jusqu'à mi-novembre. Le cri du brame tient du rugissement et du mugissement et s'entend à plusieurs kilomètres de distance. C'est aussi le terme désignant le rut chez cette espèce.
Le mâle avertit ainsi les femelles réceptives de sa présence, intimide ses concurrents potentiels et défie les autres mâles qui s'aventureraient sur son territoire. Il devient particulièrement agressif à ce moment-là. En cas de rencontre avec un autre mâle, après une phase d'intimidation, les deux adversaires vont mener un combat très violent durant lequel ils se projettent la tête en avant l'un contre l'autre dans le but de déséquilibrer l'adversaire. Ces combats peuvent conduire à l'abandon ou à des blessures assez graves voire la mort par épuisement des deux cerfs s'ils restent coincés par leurs bois emmêlés : seuls des mâles de puissance et de ramure comparables s'affrontent de la sorte.
Une étude a montré que la testostérone joue un rôle majeur dans le comportement agressif et reproducteur (uniquement en saison du rut) des mâles[17]. Les cerfs mâles expérimentalement castrés perdent leur agressivité ainsi que les comportements typiques des mâles de cette espèce, ce qui les fait descendre en termes de statut social dans le groupe. Si les cerfs castrés reçoivent de la testostérone en décembre, ils retrouvent tous les comportements de rut en quelques semaines, y compris l'agressivité envers les autres mâles. Si la testostérone leur est fournie en avril et juin, elle n'induit pas de comportement immédiat de rut mais l'animal retrouve son niveau d'agressivité sociale, puis en automne le comportement normal de rut, au moment normal du rut.
L'injection de testostérone à des cerfs normaux, quelle que soit la saison, n'induit pas une seconde période d'activité reproductrice, par contre les animaux montrent de nets changements d'agressivité. Ce n'est pas l'odeur de l’œstrus ou la présence des femelles qui rendent les mâles agressifs. Cette agressivité est également constatée chez des mâles élevés en enclos loin des femelles[14].
Le cycle des bois du cerf est également contrôlé par le niveau de testostérone. Ceci explique que les expériences visant à tester les effets d'injection de testostérone ont aussi eu des effets sur les bois, qui ont eux-mêmes affecté certains comportements. Les effets directs de la testostérone ne sont certains que quand ils se manifestent lorsque le traitement hormonal n'induit pas de changements dans les bois.
Le mâle régnant sur une harde de femelles surveillera de façon intensive les différents individus de la troupe en vue de l'accouplement, car les femelles ne restent sexuellement réceptives qu'une seule journée durant l'année. Lorsque le « maître de harde » a éliminé ses concurrents, polygame, il couvre les 10 à 30 biches de son harem au fur et à mesure de leurs chaleurs. Une biche qui est stérile est appelée « bréhaigne ».
De jeunes mâles profitent parfois des combats entre deux mâles plus forts pour saillir une biche « en chaleur », ce qui contribue à la diversité génétique de la harde.
La temporalité du cycle sexuel et l'œstrus de la femelle sont principalement sous le contrôle d'une hormone majeure, la mélatonine[18], dont le cycle est contrôlé par la durée du jour (ex. : l'injection de mélatonine à de jeunes biches avance le moment de leur première ovulation[18]). La mélatonine influe elle-même sur le cycle de sécrétion d'une autre hormone, la progestérone[19]. La production de cette hormone, selon les mesures faites dans les échantillons de sang veineux jugulaire de cerfs d'élevage, varie selon le statut de dominance de la femelle, mais non selon son poids corporel, ni selon la taille de l'enclos d'élevage. Mais le taux de progestérone des biches chute (plus que divisé par deux) après de longues périodes de temps chaud et sec. Ces observations faites sur des animaux d'élevages, nourris à satiété, peuvent ne pas différer de ce qui se passe dans la nature.
La production de progestérone et la lutéine jouent un rôle dans le caractère saisonnier du cycle ovarien et les dates de l'œstrus (qui peut être plus tardif chez les biches plus jeunes, avec une saison de reproduction potentielle plus courte que pour les biches adultes[20]. Dans les élevages, le stress des animaux (jeunes notamment) semble pouvoir affecter la puberté et éventuellement provoquer des aberrations du cycle ovarien, et expliquer certains échecs de reproduction[20]. Les dates de fécondabilité peuvent varier selon le génotype (animaux provenant de l'est ou de l'ouest de l'Europe par exemple[20]).
Des expériences utilisant des implants diffusant de la mélatonine montrent qu'on peut ainsi modifier (avancer) les dates d'œstrus et par suite de fécondation et mise bas, mais non sans risque d'élevage plus difficile des jeunes si le climat leur est défavorable[21].
Gestation
La gestation du cerf élaphe était réputée durer huit mois, mais des études récentes ont montré que la durée de gestation est en fait très variable, dépendant notamment de divers facteurs environnementaux. Cette variabilité favorise probablement la survie du nouveau-né.
Une hypothèse était qu'il existe néanmoins une synchronisation (hormonale) des œstrus[22], mais aussi des naissances au sein de chaque troupeau (les biches concevant tôt et tard en saison de reproduction ayant des périodes de gestation respectivement plus longues et plus courtes). Deux études au moins ont porté sur la date de conception et la durée de gestation et ont permis de tester et confirmer cette hypothèse (pour 393 biches ayant naturellement mis bas dans deux troupeaux, pour l'une de ces deux études, et pour 91 biches dont l'œstrus et la conception ont été artificiellement déclenchés dans une plage de quatre semaines durant la période de rut naturel). Ces études ont montré que l'âge des biches gestantes intervenait aussi sur la durée de gestation, mais l'analyse des données soutient l'hypothèse, montrant que pour chaque différence de dix jours de la date de conception il y avait un changement dans la longueur de gestation de 01/09 à 04/09 jours. Les mécanismes physiologiques précis tendant à synchroniser les naissances n'ont pas encore été élucidés. Une possibilité est un effet combiné de la mélatonine (contrôlée par la durée du jour/photopériode) et d'autres interactions hormonales[23] entre individus modifiant l'induction de la parturition, ou un contrôle nutritionnel[24],[25] lié à l'âge du fœtus aux derniers stades de la grossesse[26].
Reproduction artificielle
L'insémination artificielle avec du sperme congelé est utilisée, avec des résultats mitigés[27].
La fécondation in vitro[28], et le transfert d'embryon a aussi été utilisé chez le cerf[29], pour l'élevage, ou éventuellement la sauvegarde d'espèces de cerfs ou de sous-espèces menacées.
L'ovulation multiple permet de produire plusieurs embryons[30].
Diverses méthodes (par l'observation, des analyses, ou suivi électronique) de détection de l'œstrus ont été mises au point[31]
Génétique de l'espèce et reproduction
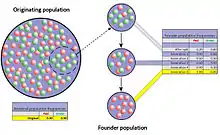
Si le noyau fondateur a une faible diversité génétique, les populations qui en découleront voient leur risque de dérive génétique ou de certaines mal-adaptations augmenter.
De manière générale, les cervidés forment un assemblage complexe de taxons avec une grande diversité morphologique, physiologique, écologique et de répartition géographique ou d'habitats (de la forêt dense à la lande écossaise ouverte). Les données scientifiques disponibles sur la génétique des cervidés sont en partie biaisées par le fait qu'elles ont surtout porté sur les espèces d'intérêt commercial (animaux d'élevages) ou jugées les moins difficiles à élever en enclos (Cervus elaphus scoticus, Cervus elaphus hippelaphus et leurs hybrides ; naturellement les plus grégaires) ; En outre, la littérature scientifique ne semble pas avoir présenté toutes les stratégies de reproduction des cervidés, qui vont de modèles très saisonniers à des naissances complètement a-saisonnières. Les espèces à stratégie K qui produisent un jeune par an durant une dizaine d'années sont de mieux en mieux connues, alors qu'on en sait peu sur les stratégies K d'autres espèces, généralement plus petites, solitaires et souvent très territoriales. Les cervidés réellement adaptés à la forêt sont souvent caractérisés par une forte fécondité (mise bas de deux jeunes) compensée par une durée de vie plus courte (<10 ans). Dans ce groupe figurent de nombreux taxons menacés de cervidés qui seraient donc probablement plus difficiles à élever faute de données sur les facteurs environnementaux influençant la gestation ou le succès de reproduction.
Quelques études ont porté sur l'analyse génétique de populations de cerfs élaphes sauvages (analyses d'ADN[32]), les croisements d'espèces différentes de cerfs (issus de zoos ou d'élevage). Elles apportent notamment des compléments d'information sur les liens entre génomes et les cycles de la reproduction[33].
D'autres études, comme en Bavière (sud de l'Allemagne) ont montré que l'espèce a en Europe de l'Ouest, comme en Amérique du Nord, connu un goulet d'étranglement génétique à la suite d'une forte réduction de population (au XVIIIe siècle en France, au XIXe siècle en Amérique du Nord) en raison d'une sur-chasse. L'espèce a depuis localement démographiquement récupéré, mais les populations échangent plus difficilement leurs gènes en raison d'une fragmentation nouvelle et croissante des habitats. Ainsi, en Bavière, l'analyse génétique d'échantillons provenant de neuf populations de cerfs et de deux populations adjacentes (Thuringe et République tchèque) a révélé que 91,6 % de ces cerfs bavarois descendent d'un même individu, et les études de corrélation entre les matrices de distance génétique et de distance géographique ne montrent pas de preuve d'isolement par la distance. Des indices de dérive génétique sont nettement présents chez plusieurs sous-populations et la diversité génétique de toutes les populations est faible, ce qui est considéré comme un facteur de vulnérabilité par les généticiens[34]. Des seuils critiques de consanguinité sont déjà dépassés dans certaines populations. Par exemple, un groupe d'une cinquantaine de cerfs du Schleswig-Holstein (Allemagne du nord) présentait un nombre anormal d'individus brachygnathes (Brachygnathie : anomalie morphologiques caractérisée par une mâchoire inférieure trop courte[35], souvent associée chez les mammifères à une dépression de consanguinité et à un retard de développement). Une analyse génétique a porté sur 18 de ces cerfs[36]. Elle a montré une très faible diversité génétique, comparativement aux individus d'autres populations européennes ou proches dont les cerfs étudiés ont dérivé depuis 130 ans[Quand ?] environ). Le noyau « originel » de cette cinquantaine de cerfs a été estimée à sept individus seulement. L'augmentation de la consanguinité correspondait dans ce cas à une perte d'hétérozygotie de 7 % pour chaque génération, valeur sept fois plus élevée que le seuil théorique au-delà duquel la sélection naturelle ne peut plus contrecarrer la fixation des allèles délétères dans le pool génétique[36]. Selon les auteurs de cette étude, cette population a un besoin urgent de gènes neufs provenant d'autres populations pour surmonter les effets négatifs de la dérive aléatoire et de la consanguinité[36].
Les analyses génétiques faites en Europe montrent que le pool génétique de l'espèce a subi maintes influences anthropogéniques. Le génome de l'espèce porte les traces des effets de l'élevage des animaux en enclos (Les populations vivant en enclos sont loin des valeurs attendues selon le modèle « isolation by distance », mais sans réduction très forte de la diversité génétique[37]), de la chasse sélective qui a favorisé les grands trophées (une corrélation a été trouvée entre certains génotypes d'alloenzymes et la morphologie des ramures, et la chasse sélective a augmenté la fréquence de ces allèles dans la direction attendue[37]), de la translocation de cerfs pour améliorer les trophées, ou de la fragmentation écologique des habitats (dont les impacts sur le pool génétique des cerfs européens restent difficiles à évaluer, faute d'état-zéro et faute d'information sur l'histoire des populations de cervidés[37].
Des phénomènes d'hybridation, d'introgression génétique[38] et de « pollution génétique » existent aussi : les études génétiques et moléculaires montrent que là où - comme en Écosse[38], dans le Nord-ouest de l'Angleterre[39] ou en Irlande - des cerfs exotiques (Cerf sika) ont été introduits du Japon, dès 1860 à partir d'individus provenant de la sous-espèce de Kyushu[40] et mis en contact avec les populations autochtones de Cerf élaphe, des croisements ont eu lieu (hybrides signalés dès 1920 en Angleterre[39]), et des translocations ultérieures d'individus ou de petites populations ont encore pu compliquer la génétique des populations[41].
Les origines de la population actuelle du Cerf élaphe en Irlande, d'après les données génétiques disponibles restent « obscures ». Les outils de la biologie moléculaire sont nécessaires pour identifier le degré d'hybridation car les apparences phénotypiques (pelage et les proportions du corps) peuvent être trompeuses. Les zones où les Cerfs élaphes et sika sont sympatriques doivent encore être évaluées pour le niveau et l'ampleur réelle de l'hybridation et le cas échéant gérées pour protéger l'intégrité génétique des populations de cerfs élaphes[41]. La situation est compliquée par le fait que certaines sous-espèces de Sika sont morphologiquement très proches d'hybrides du Cerf sika et du cerf élaphe[39]. De plus, le Cerf sika (dont les individus importés au Royaume-Uni) avait déjà lui-même subi une dérive génétique liée à la fragmentation de ses populations au Japon au XIXe siècle ou avant[40]. Localement, sur des territoires restreints, l'hybridation est très avancée. Par exemple sur la presqu'île écossaise de Kintyre, 43 % des 735 cerfs génétiquement testés étaient des hybrides alors que les deux espèces ne sont en contact que depuis 120 ans[Quand ?][42], ce qui laisse penser que le taux d'hybridation pourrait fortement augmenter dans les décennies à venir dans toute l'Angleterre là où les deux espèces sont présentes.
La mise bas (vêlage)
Dans la nature, la mise bas semble s'effectuer discrètement, un peu à l'écart du groupe, dans une zone retirée, non enherbée, à l'abri du dérangement.
Dans les élevages (fréquents en Nouvelle-Zélande), la mortalité des nouveau-nés est importante (10 % des naissances) et plus importante chez les jeunes biches primipares que chez des adultes multipares[43]. Une étude a montré que les biches d'un an maintenues en enclos montrent un comportement de stress (marche le long des clôtures) plus fréquents (43 % des observations à cette période) dans les 24 h avant la naissance[43]. Ce comportement pourrait aussi correspondre à une volonté de la biche de s'isoler, car les biches sont souvent observées isolées (à plus de 20 m des autres individus du troupeau) durant les deux jours précédant la parturition (ce comportement est aussi courant chez les adultes que chez les jeunes biches primipares).
Au moment de la parturition dans l'enclos d'élevage - lors de cette étude - 44 % des biches adultes et 60 % des jeunes biches d'un an ont eu des comportements d'interférence avec d'autres biches (remarque : les autres biches du troupeau qui ont interféré avec celles qui mettaient bas étaient plus souvent de jeunes biches d'un an)[43]. Lors de cette étude, les faons nés des biches primipares ont commencé à téter leur mère en moyenne 44 min après la naissance (c'est 11 minutes de plus que dans le cas des faons nés de biches adultes)[43].
Cette étude laisse penser que les jeunes biches inexpérimentées ont plus de difficulté à exprimer un comportement naturel de recherche d'isolement et une moindre aptitude maternelle, peut-être liée à une anxiété accrue (indiquée par la marche le long des clôtures de l'enclos), et peut être augmentée par les interférences acceptées d'autres biches au moment de la mise bas. Ceci, ainsi que le délai plus long entre la naissance et la première tétée pourrait augmenter le risque de mortalité des faons nouveau-nés[43]. De même, les faons ont l'instinct de se cacher dès après la naissance. Dans un enclos d'élevage de cerfs, si au moment de la mise bas, des caches de type fossé, hautes herbes ou amas de branches de pins sont disponibles comme alternatives à la prairie plate et ouverte, les biches et faons les choisissent et les réutilisent, plus souvent s'il s'agit d'un terrain aux herbes courtes, alors que le faon se cache de manière plus aléatoire si le terrain est constitué de hautes herbes[44] ; Dans les élevages on observe aussi régulièrement des faons tentant de passer au travers de la clôture (14,6 % des faons, que les herbes soient hautes ou non, dans l'étude précédemment citée)[44]. En élevage, que le terrain soit riche en caches ou non, les interactions entre le faon et des biches autres que sa mère sont fréquentes (les faons approchés ou suivis, reniflés, léchés ou ayant reçu des coups ont été observés respectivement dans 25, 34, 5 et 14 % des cas[44]. Divers auteurs estiment que dans les élevages, le vêlage et l'élevage des faons devraient être faits dans des enclos où l'herbe est haute ou avec des cachettes disponibles pour les animaux. D'autres auteurs ou les mêmes estiment qu'il est aussi utile d'offrir des caches aux biches au moment du vêlage[44].
Alimentation
Le cerf élaphe est un herbivore et un ruminant, mais comme tous les mammifères, il est d'abord nourri, après la naissance du lait de sa mère, dont les teneurs en sels minéraux[45] s'adapteront au poids du faon, jusqu'au sevrage.
Au sein de la classe des herbivores, le cerf est dit intermediate feeder, c'est-à-dire qu'il est très sélectif dans son alimentation, tout en s'adaptant toujours à la végétation qu'il a à disposition.
Au sein des massifs résineux de montagne, le cerf préfère le sapin (Abies alba) à l'épicéa (Picea abies). Au sein des massifs feuillus de plaine, son comportement alimentaire varie selon la saison et les plantes disponibles :
- de la sortie de l'hiver jusqu'à l'automne, il se nourrit de bois (il mange les bourgeons et les jeunes pousses des arbres et arbustes, sauf les épineux qu'il évite), de graminées, lierre, ronce et autres plantes herbacées dont il consomme parfois les fleurs. Toutefois, les forêts de plaine étant fréquemment entourées de cultures, il va souvent se nourrir de maïs ou de colza et peut alors être directement exposé aux pesticides.
En fin de saison, il consomme également des fruits (pommes, poires).
- en hiver, il se nourrit de bois, feuilles de ronces, feuilles mortes et de ce qui reste à sa disposition. Son régime varie selon la présence de neige et selon la glandée ou fainée qui se produit sur la forêt.
Un adulte consomme en moyenne de 10 à 15 kg de végétaux frais par jour :
Déplacements et fragmentation croissante des paysages
Les cerfs et plus encore les biches sont dotés d'une musculature puissante, avec de longues pattes fines et légères terminées par des sabots serrés et pointus, adaptés à la course et aux bonds. Ils peuvent courir à 40 km/h en vitesse de croisière mais en cas de danger, les biches et certains cerfs (les plus minces) peuvent courir très vite jusqu'à 76 km/h en pointe[46][réf. non conforme]. Ils peuvent aussi faire de grands sauts (jusqu'à 2,50 mètres en hauteur et 9 mètres en longueur pour les biches contre 3 mètres de haut à 11 mètres de long pour les cerfs[47], il s'agit là de records mesurés sur des cerfs sauvages), et ils ne sont pas arrêtés par l'eau. Par contre, les gros cerfs mâles plus âgés courent moins vite que les biches et les jeunes cerfs, mais ils sont plus forts au combat (du moins quand ils ont leurs bois). Les animaux âgés, les très jeunes faons ou encore les animaux en moins bonne santé ne sont pas très rapides à la course, et constituent généralement des proies idéales pour les prédateurs. En l'absence de leurs prédateurs naturels (les loups), dans certaines régions d’Europe (notamment en France), les cerfs se déplacent moins et sont moins musclés (quels que soient leur âge, leur sexe ou leur poids). De plus, ils sont effrayés par des obstacles ou objets qu'ils ne connaissent pas (simples drapeaux sur corde tendue par exemple). On peut supposer qu'autrefois, en présence de grands prédateurs tels que le tigre à dents de sabre, le lion des cavernes ou de prédateurs tels que le loup et la hyène et le lynx, les cerfs se déplaçaient sur de plus grandes distances et saisonnièrement, mais avec la fragmentation des massifs forestiers et milieux naturels, ils tendent à se sédentariser.
Si leurs prédateurs sauvages ont disparu, comme beaucoup d'animaux chassés depuis longtemps par l'homme ou d'autres prédateurs, les cerfs continuent à se déplacer avec prudence, surtout dans les lieux qu'ils ne connaissent pas encore.
Face à une fragmentation forestière croissante, et pour notamment mieux concevoir ou protéger les corridors biologiques ou les écoducs construits pour leur permettre de mieux circuler dans le territoire, des éthologues cherchent à mieux comprendre leurs déplacements.
- Par exemple, huit groupes de six cerfs issus d'élevage ont été placés dans des couloirs de différentes formes (droits ou courbés) et largeur (0,5 m ou 1,5 m de large) et plus ou moins éclairés[48]. Les réactions de ces animaux ont été observées dans des circuits avec huit combinaisons de traitements. Ils sont entrés plus vite dans les couloirs les plus larges (après 2,7 secondes en moyenne, contre 6,9 pour les couloirs plus étroits). Ils sont entrés plus vite aussi dans les couloirs droits que dans ceux incurvés (p < 0,05). Et, ils couraient plus vite dans les couloirs larges qu'étroits (1,3 m/s contre 1,0 m/s en moyenne ; p < 0,01). Dans tous les cas, ils se déplacent plus volontiers en groupe que les uns après les autres en file[48]. La forme du parcours ou l'intensité lumineuse n'ont pas beaucoup modifié la vitesse ou l'espacement entre les cerfs. Plus l'expérience était répétée (habituation), plus les cerfs couraient vite, avec une forte augmentation de la vitesse dès la seconde expérience, mais peu de changement par la suite[48].
- In situ, en Espagne, en posant des barrières artificielles modestes (clôtures telles que l'on en trouve souvent posées pour protéger la végétation de l'impact des cervidés et sangliers), on a montré que ces clôtures empêchent non seulement les cerfs de pénétrer la zone clôturée elle-même, mais qu'elles diminuent aussi l'utilisation des zones adjacentes par les animaux, au point d'interrompre parfois des voies de déplacement antérieurement majeures. En Espagne, des clôtures ont ainsi été placées entre les principaux domaines d'alimentation et de repos des cerfs (durant la saison des amours, de manière à étudier leurs effets éventuels sur la répartition des femelles parmi les harems). Les chercheurs ont constaté que les zones d'ombre situées dans l'axe des déplacements, à proximité des nouveaux obstacles ont été moins utilisées par les cerfs (ce qui s'est aussi traduit par des graminées plus hautes, qui indiquent une sous-exploitation des ressources de la zone par le cerf[49].
- Les déplacements forcés (fuite devant un prédateur, devant le dérangement par l'Homme ou lors de la chasse, chasse à courre notamment) sont des sources importantes de stress pour ces animaux[50].
- On a aussi étudié le stress causé par le déplacement en camion de cerfs issus d'élevage, notamment dans les conduites aux abattoirs ; par exemple, neuf groupes de six biches issues d'élevage ont été transportés sur des distances de 80, 230 ou 380 km (représentatives des pratiques commerciales courantes en Amérique du Nord)[51]. Une caméra fixée sur le plafond du camion filmait le comportement des animaux en continu et des échantillons sanguins ont été prélevés avant le transport et à l'arrivée avant l'abattage. L'état des carcasses a aussi été évalué (mesure du pH et comptage des ecchymoses. Les efforts d'ajustements d'équilibre (mouvement rapides des pieds et nombre d'impacts) étaient, comme on pouvait s'y attendre, plus nombreux chez les animaux plus grands, au début du voyage et sur les routes raides ou sinueuses[51].
Le nombre d'ecchymoses a été jugé plutôt faible, et celles qui ont été observées sur les membres postérieurs et au niveau des vertèbres augmentaient avec la distance de transport. Les morsures ont représenté 95 % de tous les comportements agonistiques observés via la caméra durant le transport, et les animaux les plus lourds étaient ceux qui commençaient la plupart des comportements agonistiques, en les orientant généralement vers les petits animaux. Durant les deux tiers des voyages, un ou plusieurs animaux se sont couchés au sol. Ces biches s'allongeaient alors sur le fond du camion en moyenne après 60 min de transport[51]. Les analyses sanguines ont montré que le stress induit par le parcours avant l'abattage (transport compris) se traduit par d'importantes modifications hormonales et enzymatiques du sang, en particulier par une augmentation de la créatine kinase (CK), de l'aspartate aminotransférase (AST), du glucose sanguin, de la calcémie, du cortisol sanguin, du lactate déshydrogénase total (LDH), LDH3, LDH4, LDH5 et par une chute du magnésium sanguin. Le temps de trajet n'a pas d'effet sur le comportement, la qualité des carcasses (ecchymoses) ou le pH ultime, mais est en soi un facteur de stress (traduit par une augmentation de CK, LDH totale, et LDH5 LDH4 proportionnelle à la distance parcourue par le camion)[51].
Suivi et dynamique des populations
À la suite d'une chasse intensive, les populations de grands cervidés avaient fortement régressé en Europe de l'Ouest et du Sud et en Chine dès la fin du Moyen Âge. À la fin du XIXe siècle, le cerf élaphe avait disparu de la plupart des massifs forestiers de l'Europe de l'Ouest.
Avec l'organisation et la règlementation progressive de la chasse depuis le XIXe siècle (réintroductions, élevages, nourrissage, plans de chasse…) de nombreuses populations se sont localement reconstituées et continuent parfois à augmenter[52] à partir d'individus réintroduits, mais avec un appauvrissement génétique probablement important par rapport au patrimoine génétique des populations préhistoriques.
Le cerf est un des premiers animaux terrestres européens considérés non menacés à avoir fait l'objet d'études ADN (par exemple sur le massif de Saint-Hubert en Belgique[53]), permettant d'assigner avec certitude des mues à une série ou à un trophée, d'apparier les parties d’animal qui auraient fait l'objet de trafic ou braconnage. On a aussi pu démontrer l'existence en Région wallonne de trois sous-populations génétiquement isolées en raison des barrières autoroutières. Un poil ou un morceau de viande crue saisis dans un restaurant ou un véhicule suffisent à déterminer l’espèce, le sexe, et, quand des études antérieures existent (en Région wallonne par exemple), la provenance de l'animal. Une analyse PCR basée sur des amorces oligonucléotidiques ciblant le gène ARNr 12S mitochondrial a été récemment mise au point pour lutter contre les fraudes de viandes ou produits carnés à base de chair de cerf élaphe (Cervus elaphus), daims (Dama dama) et chevreuil (Capreolus capreolus)[54].
Dans de nombreux pays, les populations de cerfs qui se sont reconstituées sont de plus en plus isolées par la fragmentation du paysage par les infrastructures (et parfois elles ont été maintenues dans des surfaces encloses), sur des territoires parfois restreints où elles survivent grâce à l'agrainage et parfois en surexploitant les sous-bois.
Dix-huit cerfs parmi cinquante issus d’une population du nord de l’Allemagne ont fait l'objet d'une analyse génétique[55]. Cette population était supposée écologiquement isolée des autres populations par des infrastructures routières depuis 30 à 40 ans, et de nombreux cerfs dans ce groupe sont affectés d'une malformation génétique rendant leur mâchoire inférieure plus courte d’environ 5 cm par rapport à la supérieure. L'analyse génétique a confirmé la perte de diversité génétique du groupe, avec une perte d’hétérozygotie d'environ 7 % à chaque génération. C'est sept fois plus que dans la population voisine d'où elle est originaire, et cela rend ce type de population d'autant plus vulnérables aux anomalies génétiques qu'en l'absence de grands prédateurs, les animaux sont moins mobiles, plus sensibles aux parasites et échappent aux processus de sélection naturelle.
Problèmes liés à l'augmentation des populations et/ou à la chasse

La chasse aux grands cervidés est ancienne. Bien avant l'invention du fusil, elle a été grandement facilitée par le dressage de chiens de chasse.

L'usage du cheval est un autre facteur qui a facilité la chasse des cerfs, avec notamment la chasse à courre.


Si la chasse a un temps failli faire disparaître les cerfs d'Europe et Asie, les réintroductions, l'agrainage, les apports en sel et les « plans de tir » mis en place au XXe siècle ont en Europe donné des résultats qui ont dépassé les espérances des acteurs qui ont lancé les dynamiques de gestion restauratoire de population de grands et petits cervidés.
Comme les sangliers et les chevreuils bénéficiant outre d'hivers plus doux, de compléments d'alimentation qui leur sont offerts en forêt (agrainage, cultures cynégétiques), les populations de cerfs tendent à atteindre des « records[56] » populationnels dans plusieurs pays de l'Union européenne. Outre des dégâts sylvicoles on peut craindre, en raison de l'isolement de la plupart des populations, et en l'absence de sélection par leurs prédateurs naturels carnivores (qui ont presque disparu), des problèmes éco-épidémiologiques avec diffusion de maladies favorisées par la promiscuité et l'absence de pression sélective par prédation naturelle, voire des impacts sur l'homme avec par exemple la diffusion de maladies à tiques, dont la maladie de Lyme et les pullulations de tiques[57] ou peut-être des maladies telles que la CWD.
On suppose qu'une densité trop élevée de cerf par massif ou par territoire devient - au-delà d'un certain seuil - un facteur de stress pour les cerfs eux-mêmes. On note dans les élevages, que les mâles en particulier se montrent plus agressifs entre eux, et un peu différemment (de manière moins sélective) de ce qui se passe dans la nature concernant le système de dominance[58],[14], lorsqu'ils sont confinés sur de petites surfaces. De plus, de manière générale, au moment du rut, les cerfs mâles dépensent beaucoup d'énergie, ce qui se traduit par une perte importante de poids et ils peuvent être affaiblis par leurs blessures lors des combats.
Des éthologues se sont demandé si la présence d'abris légers (naturels ou artificiels) sur leur territoire pourrait modifier leur comportement face à divers facteurs de stress. Une étude a ainsi observé les réponses comportementales de huit groupes de cinq jeunes cerfs (groupes mixtes de mâles et femelles) à diverses perturbations (stimuli visuels (escalade d'une clôture) et sonores ; ici diffusion de musique forte) sur un territoire avec ou sans abris (abris légers de type windcloth en trois sections de 4 × 1,5 m). Dans cette expérience, la présence d'abris artificiels n'a pas eu d'incidence sur la position des sujets, ni sur la fréquence ou la durée des activités observées des cerfs (comportement agressif, pâturage, comportement d'alerte). Cependant, les réponses à deux types de perturbations ont été modifiées ; les cerfs étaient plus alertes lors de présentations d'un stimulus visuel et des taux plus élevés d'agression étaient notés ensuite. En revanche, le bruit (musique) conduisait à plus d'agressivité au moment du stimulus et à un niveau d'alerte plus élevé des cerfs après le stimulus. Selon les auteurs, ces deux types de perturbations (escalade des clôtures ou musique forte) n'ont pas semblé provoquer de changements durables de comportement[59].
L'impact comportemental de la taille des enclos d'élevage a aussi été étudié. Par exemple, quatre groupes de dix cerfs issus d'élevage, âgés de 2 ans, ont été temporairement confinés (de 40 min à 2 jours) dans des enclos de 5 × 4 m ou dans des enclos deux fois plus petits (2,5 × 4 m), au printemps et en été[60]. Dans les petits enclos, les marches lentes le long des grillages et des mouvements de tête verticaux et horizontaux ont été plus fréquents (p <0,05) et ont été pratiqués par un pourcentage plus élevé des cerfs (P <0,01), et les distances entre individus étaient plus petits que dans les grands enclos[60]. L'agressivité a varié selon les saisons : les coups de tête et poursuites étaient plus fréquentes au printemps et les morsures et coups de pied plus fréquents en été, mais avec une agressivité globalement moins fréquente en été[60]. Au printemps, dans les petits enclos, il y avait moins de menaces par coup de tête, moins de coups de tête donnés par des animaux en mouvement, et moins d'activité de marche que dans de grands enclos. En été, dans les petits enclos, il y avait plus de menaces envers les arrière-trains et plus d'activité de marche que dans les enclos de grande taille. En été comme au printemps, l'agressivité des cerfs était corrélée à l'activité de marche le long des clôtures[60].
Ceci laisse penser qu'on peut minimiser l'agression interindividuelle en plaçant les cerfs d'élevage dans des enclos plutôt petits au printemps et plus grands en été. mais l'observation des distances inter-individuelles et des mouvements de tête montrent que quelle que soit la saison, les grands enclos sont préférés par les cerfs[60].
Équilibres « sylvo-cynégétiques »
À titre d'exemple, en Région wallonne, du au , ce sont 4 732 cerfs qui ont été abattus ou trouvés morts (1 401 cerfs « porteurs de bois », 3 328 cerfs « non boisés » dont 1 571 biches et bichettes et 1 757 faons ; 842 mâles, et 770 femelles, 145 indéterminés par les déclarations ; et 3 non identifiés), ce malgré une augmentation du nombre de tirs. Ces tableaux sont de nouveaux records et inquiètent les forestiers wallons alors que la répartition des tirs et mortalités entre biches, bichettes et faons semble correspondre aux objectifs (45 à 50 % de biches et bichettes pour 55 à 50 % de faons). Les statistiques montrent que depuis 1996, les chasseurs privilégient encore la survie des femelles (sauf pour 1999), de celles des faons mâles (tués à la chasse, ou retrouvés morts), le pourcentage de ces derniers a toujours été plus élevé que celui des faons femelles. Les taux de non boisés tirés à l'approche et à l'affût (soit 20 % des cerfs non boisés tués) ont varié de 0 à 81 selon les conseils cynégétiques considérés (il y en a 23 en Région wallonne, dont 17 pratiquant ce mode de chasse). Il reste difficile de juger à partir de quand on peut parler de surpopulation, d'autant que le cerf est une source importante de revenu forestier via la location des droits de chasse et que les comptages sous-estiment toujours les effectifs[61]. En France, la situation est identique : 43 744 cerfs ont été abattus en 2007 par les chasseurs (hors parcs et enclos) à la saison 2006-2007, ce qui correspond à une progression de + 5,2 % par rapport à l'année précédente, et à + 410 % par rapport à 1986 mais seulement à 74,2 % de la réalisation du plan de chasse[62],[63]. Les populations devraient donc encore augmenter[réf. souhaitée].
La grande « école du cerf » est en France la réserve nationale de Chambord : l'ancien domaine royal s'étend sur 5 343 ha de bois, d'étangs et de pâturages, derrière une muraille de 33 km. Quinze spécialistes payés par l'État (aidés de trente ouvriers agricoles) étudient 600 cerfs et 900 sangliers. Seuls des invités officiels y sont admis à chasser (à la carabine, à l'approche ou à l'affût). Ils sont toujours accompagnés de conseillers qui leur désignent les pièces à tuer. On y capture aussi, chaque année, une centaine d'animaux vivants destinés à repeupler les autres massifs français. « Chambord doit être d'abord une école vivante », souhaite Christian Mary, commissaire général du domaine[citation nécessaire]. Dès la mi-septembre, des dizaines de curieux se pressent chaque nuit dans les miradors pour observer le brame. Cinq mille écoliers s'y rendent chaque année en groupes et des visites commentées y sont organisées.
Les cerfs sont 37 000 en France dont plus des trois quarts dans la moitié du nord du pays (surtout en Champagne, Alsace et Lorraine). Leur population doublerait tous les deux ou trois ans, si la chasse était supprimée, car douze mille petits environ naissent chaque printemps. De dix à onze mille cerfs sont tués chaque année par les chasseurs. Une gestion « saine » des effectifs exigerait que, chaque année, sur cent animaux, une trentaine soit tués, dont neuf faons mâles et femelles et dix daguets et bichettes de plus d'un an. Il faudrait aussi tuer de préférence des individus déficients, comme le feraient les carnassiers dans la nature. Cependant, les chasseurs sont laissés libres : la plupart choisissent selon la quantité de viande et la beauté des bois alors que la chasse doit avoir pour premier but le contrôle et la sélection des populations.[réf. souhaitée]
Certaines sources estiment[64] que, comme le chevreuil ou le sanglier, le cerf peut causer de graves dégâts aux forêts où il est en surpopulation. Dans le parc national des Cévennes, la forêt telle qu'elle existe en serait menacée. Environ 15 000 ha sont classés en zone interdite à la chasse, ce qui, en l'absence de prédateurs carnivores sauvages, aurait entraîné une multiplication anormale des cervidés[65]. De tels dommages pourraient peut-être être évités en réintroduisant ou acceptant le loup dans le parc des Cévennes.
Un problème identique avait été identifié dans le parc américain de Yellowstone : une surpopulation de wapitis mangeant les pousses d'arbres entraînait une déforestation. La réintroduction du loup, prédateur de wapitis, a permis d'en contrôler la population et ainsi d'enrayer la menace sur la forêt. La réintroduction du loup a également permis de restaurer l'équilibre naturel de l'écosystème du parc[66], perturbé depuis l'extermination du loup par l'homme dans cette zone, dans les années 1950. Cependant, les caractéristiques (taille du parc, faible nombre de sentiers et de routes…) du parc de Yellowstone limitent la comparaison.
Aspects sanitaires
Concernant la santé des cerfs et de leurs populations

Les populations de cervidés (et d'autres ongulés sauvages) « peuvent avoir un rôle important dans le maintien de populations de tiques, et aussi en tant que réservoirs de germes pathogènes ». À titre d'exemple, on a trouvé en Espagne dix espèces différentes de tique parasitant le cerf élaphe (dont principalement Hyalomma marginatum (63,7 %), Rhipicephalus annulatus (en) (7,9 %) et Rhipicephalus bursa (de) (7,5 %), avec des prévalences géographiques très variables. Et en moyenne, les cerfs portaient près de quatorze tiques par individu, soit à peu près autant que les sangliers (13,6 par sanglier), mais, bien que partageant les mêmes territoires, seules cinq des treize espèces de tiques ixodidées trouvées parasitaient à la fois le cerf et le sanglier, ce qui laisse penser que les tiques ont des préférences en matière d'hôtes[67].
Le cerf élaphe est par ailleurs naturellement sensible à certaines maladies dont :
- toxoplasmose, par exemple détectée chez 15,6 % de 441 cerfs élaphes testés en Espagne[68], sans différence de séroprévalence selon le sexe, l'âge ou la gestion des domaines de chasse (ouvert ou clôturé). C'est en Espagne la plus forte séroprévalence enregistrée chez les ruminants sauvages pour T. gondii (qui était de 24 % pour 79 daims (Dama dama), 21,8 % chez 33 chevreuils (Capreolus capreolus), 33,3 % pour trois bouquetins espagnols (Capra pyrenaica), 20 % de dix chamois (Rupicapra pyrenaica), 10 % de dix mouflons (Ammotragus lervia), et 14,8 % de 27 mouflons (Ovis ammon), ce qui suggère que la consommation de viande de cerf mal cuite, tout comme la manipulation des viandes et carcasses de gibier sauvage, devrait être considérée comme facteur de risque d'infection pour les humains)[68] ;
- babésiose[69] ;
- paratuberculose : les faons ou cerfs adultes semblent développer peu de symptômes cliniques face à la paratuberculose, mais sont infectés par l'inoculation par la bouche d'une forte dose de microbes[70] ;
- CWD : on note aux États-Unis l'apparition récente d'une maladie émergente ; Encéphalopathie mortelle à prions, proche de la maladie de la vache folle, dite CWD, qui a pris une extension rapide dans les années 1990.
Les cerfs, bien moins que les animaux carnivores, peuvent aussi être exposés à une bioaccumulation de métaux lourds pouvant affecter leur santé. Une étude a porté sur des cerfs de la vallée d'Alcudia (es) et de la Sierra Madrona (es) (sud de l'Espagne) exploitée pour les métaux (mines) il y a plus de 2 100 ans par les Romains puis périodiquement et au XXe siècle jusqu'à la fin des années 1900[71]. Chez 168 cerfs élaphes et 58 sangliers, des analyses de plomb, zinc, cuivre, cadmium, arsenic et sélénium ont été faites sur des échantillons de foie et d'os (métacarpe)[71]. La peroxydation lipidique, le glutathion total, le glutathion oxydase (GSH), la superoxyde dismutase (SOD), le glutathion peroxydase (GPX) et composition en acides gras ont aussi été étudiés dans les foies des cerfs[71]. Les teneurs des foies et os de cerf et sanglier étaient effectivement plus élevés dans le secteur minier que dans la zone de contrôle. Les sangliers en accumulaient plus que le cerf ; toujours sous le seuil associé aux signes extérieurs du saturnisme animal, mais avec un métabolisme très affecté, avec par exemple 39 % de GSH total en moins que dans la zone de contrôle[71]. Le taux d'acide docosahexaénoïque des foies de cerf élaphe de la région minière était de 16 % inférieur à celui de la zone de contrôle. Ceci est aussi attribuable aux taux de Cu, Cd et Se également plus élevés dans les foies des cerfs élaphes de la zone minière que dans la zone témoin[71]. Il a été proposé en Écosse de remplacer les balles de plomb par des balles alternatives moins toxiques lors des battues au cerf, pour limiter les risques de saturnisme induit par les munitions en plomb.[72]
Ces métaux ou d'autres toxiques, ainsi que le stress pourraient rendre les cerfs touchés plus sensibles aux maladies en zones de surdensité, à cause de la conjonction d'une baisse d'immunité avec la promiscuité, à quoi se surajoute l'impossibilité fréquente de changer de territoire (car leur environnement est morcelé et manque de corridors biologiques et d'écoducs adaptés aux grands mammifères herbivores). Les zones de forte densité de cervidés correspondent d'ailleurs souvent à des zones de pullulation de tiques, acariens piqueurs-suceurs connus comme vecteurs de nombreuses maladies (dont des zoonoses). (Tuberculose, babésiose (maladie à laquelle le cerf Sika semble mieux résister que le cerf élaphe[69]), souvent transmise en Europe par la tique Ixodes ricinus qui semble en extension).
De même, on a montré (2007) avec les Wapitis de Yellowstone que l'agrainage a aussi des effets négatifs : attirer un nombre important d'individus au même endroit, notamment en hiver, est aussi un facteur exacerbant la transmission de parasites, dont via les excréments qui contaminent la nourriture, ou en favorisant la promiscuité d'individus sains avec d'autres malades et contagieux[73]. L'offre en pierre à sel semble utile dans un environnement fermé où des carences sont possibles, d'autant que les cervidés semblent capables de consommer les sels selon leurs besoins[74], mais cette offre en sels, comme en aliments appétents ou d'autres techniques (clôtures…) visant à fixer les cerfs localement dans un massif ou un territoire de chasse peuvent avoir des effets de consanguinité et sur la santé des cerfs eux-mêmes, mais aussi sur des animaux de fermes proches ou pour l'Homme quand les maladies sont des zoonoses[73].
Concernant la santé des consommateurs

Les cerfs peuvent bioaccumuler des métaux (cadmium par exemple[75] ou mercure, avec par exemple jusqu'à 3.71 mg de mercure par kg de muscle, pour la période 2002-2008 en Croatie[76]), minéraux (fluorures par exemple[77]) et des radionucléides (à la suite des retombées de la catastrophe de Tchernobyl par exemple, ou lorsqu'il pâture en zone granitique[78]), notamment quand ils sont jeunes au moment de la rapide croissance du squelette. Les métaux sont retrouvés dans le foie, le rein, le cerveau, la viande (et les os pour le plomb). Les taux de métaux bioaccumulés varient selon les organes, les animaux et leur provenance.
Il peut arriver que les balles de plomb, notamment quand elles éclatent au contact d'un os, ou quand le cœur a continué à battre quelques instants après la pénétration de la balle, contaminent la viande. En théorie, le boucher doit supprimer la viande périphérique au trou de pénétration de la balle. Il existe pour la chasse au grand gibier des munitions sans plomb dites non toxiques.
Localement, en mangeant dans des champs récemment traités par des pesticides, ou en les traversant, ils peuvent aussi se contaminer par ces produits.
En raison de la perte annuelle des bois, le besoin en minéraux des mâles est comparativement plus important que chez d'autres animaux. Les biches ont des besoins importants durant la gestation et l'allaitement, même si comme beaucoup de grands herbivores, elles ne produisent qu'un jeune par an.
Comme les troupeaux sauvages sont difficiles à échantillonner, une étude a porté sur l'évaluation des teneurs corporelles en Césium 137 à partir de l'analyse des excréments[78].
La dangerosité pour l'homme des prions pathogènes infectieux de la CWD est encore discutée, mais des précautions similaires à celles prises pour limiter les risques dans le cas de la vache folle sont recommandées. Le velours des bois, riche en sang et hormone de croissance a été utilisé pour produire des compléments alimentaires aux États-Unis[79] ou des médicaments, de longue date dans les pays asiatiques et semble-t-il dans la Rome antique, et autrefois en Europe, Asie et Amérique du Nord ou plus récemment en URSS (dans la pharmacopée officielle). Il aurait notamment été prescrit contre les douleurs articulaires, des problèmes rénaux et hépatiques, l'ulcère à l'estomac, la goutte, les maux de tête, l'eczéma, les frissons. Les Chinois lui attribuaient, prescrit en poudres, pilules, extraits, teintures et pommades de nombreuses vertus dont pour renforcer les os et tendons, pour nourrir le sang, réduire les œdèmes ou pour lutter contre l'impuissance[79]. La Chine produit près de 100 tonnes par an de bois de velours séchés[79], c'est une ressource que la Nouvelle-Zélande qui dispose de nombreux élevages de cerfs veut exploiter[80] et la Russie en exporte vers le Japon, Singapour, Hong Kong et en Thaïlande, mais l'Asie apprécie les bois importés d'Amérique du Nord, car plus grands et considérés comme plus actifs que ceux d’andouillers couramment disponibles[79]. Or, en Amérique du Nord, le velours du cerf peut contenir des prions (CWD).
Symbolique
Le dieu cornu de l'Antiquité
.jpg.webp)
Une figure christique au Moyen Âge

Au Moyen Âge, le cerf est un animal à la charge symbolique particulièrement forte[81]. À l'égal de l'ours ou du lion, il fait partie des royautés animales. Des auteurs comme Bède le vénérable ou Raban Maur en font l'image du chrétien, de l'homme innocent, pur et saint. La légende de l'invention des reliques de saint Denis, trouvées par Dagobert sur les indications d'un cerf envoyé par la Providence renforce cette idée. Les hagiographes de saint Hubert ou de saint Eustache l'associent plus particulièrement au Christ, apparu en croix à ces deux saints entre les bois d'un cerf. Divers parallèles sont établis en ce sens par les lettrés du Moyen Âge. Les livres de vénerie insistent ainsi sur le fait que le cerf est un animal destiné à être sacrifié au terme d'un rituel précis, comme le Christ a été rituellement sacrifié. De même, les bois du cerf, repoussant chaque année après être tombés, apparaissent comme des images de la résurrection. Le cerf à la robe d'un blanc immaculé devient un véritable symbole christique[82].
Cette association explique le succès de cet animal auprès des rois souhaitant démontrer leur piété. Richard II d'Angleterre choisit ainsi le cerf blanc couché sur une prairie que l'on voit au dos du diptyque Wilton comme emblème personnel. C'est néanmoins dans les rois de France du XVe siècle que le cerf trouve les plus fidèles dévots. Si le cerf fait discrètement partie du vocabulaire traditionnel de la monarchie, c'est Charles VI qui, le premier, donne à cet animal une réelle importance dans le bestiaire royal, sous la forme du cerf ailé (parfois appelé cerf volant ou cerf de justice). Philippe de Mézières introduit ainsi ce thème dans le Songe du viel pèlerin, décrivant le roi comme un « noble cerf ailé », image du Christ sur Terre. Cette comparaison aura un immense succès pendant tout le XVe siècle. Son fils Charles VII, reprend cette symbolique pour affirmer sa légitimité et sa filiation, faisant souvent représenter ses armoiries tenues par deux cerfs blancs ailés portant une couronne autour du cou. Louis XI, peu enclin à la pompe symbolique et à la célébration allégorique de son pouvoir, délaisse quelque peu les cerfs qui seront à nouveau à l'honneur sous Charles VIII et Louis XII. Celui-ci est le dernier à être chanté comme le cerf de France, François Ier et successeurs abandonnant totalement ce symbole. Pendant toute cette période, les cerfs ailés font partie du répertoire iconographique de la monarchie, abondant dans les tapisseries, les manuscrits et les décors monumentaux. Certains princes, comme Pierre II de Bourbon suivent la mode et adoptent également cet animal.
Arts martiaux
Le cerf, dans l'art martial chinois du jeu des 5 animaux, correspond à plusieurs figures censées imiter cet animal.
Représentations dans la peinture
- Les Cerfs de Rosa Bonheur

Biche et faon dans un sous-bois, 1868
Collection privée, Vente 2007
Le Monarque du troupeau, 1868
Collection privée, Vente 2009_-_The_Stag_-_FA000258_-_Brighton_Museum_%5E_Art_Gallery.jpg.webp)
Le Cerf, 1875
Musée et galerie d'art de Brighton
Trois cerfs dans un bois, 1875
Collection privée, Vente 2005
Cerf vigilant, 1883
Collection privée, Vente 2007.jpg.webp)
Cerf et biche dans un paysage enneigé, 1883
Collection privée, Vente 2013
Un cerf, 1893
Galerie nationale d'Irlande
Notes et références
- . Pour une liste des autres sous-espèces de cerf, voir le genre Cervus.
- Larmière ou larmier, Dictionnaire de la langue françoise, ancienne et moderne.
- (en) Encyclopedia of Quaternary Science p. 3158-3175 DOI:10.1016/B0-44-452747-8/00262-3.
- (en) Caloi, Lucia et Palombo, Mari Rita, « Functional aspects and ecologicalimplications in Pleistocene endemic cervids of Sardinia, Sicily and Crete », Geobios, vol. 28, no 2, , p. 247-258 (DOI 10.1016/S0016-6995(95)80231-2, lire en ligne).
- (en) Anna Stankovic, Karolina Doan, Paweł Mackiewicz, Bogdan Ridush, Mateusz Baca, Robert Gromadka, Paweł Socha, Piotr Weglenski, h, Adam Nadachowski et Krzysztof Stefaniak, First ancient DNA sequences of the Late Pleistocene red deer (Cervus elaphus) from the Crimea, Ukraine ; Quaternary International ; DOI:10.1016/j.quaint.2011.05.023, mis en ligne le .
- (en) Ludt, Christian J., Schroeder, Wolf, Rottmann, Oswald et Kuehn, Ralph, « Mitochondrial DNA phylogeography of red deer (Cervus elaphus) », Molecular Phylogenetics and Evolution, vol. 31, no 3, , p. 1064-1083 (DOI 10.1016/j.ympev.2003.10.003, lire en ligne)
- Moulin de Poyaller.
- Bois de cerf.
- Bois de cerf.
- La fiche du cerf élaphe, sur le site de l'ONCFS, consultée en 2017.
- « Les espèces animales les plus menacées par les incendies », sur Geo.fr,
- Schéma départemental de gestion cynégétique de la Meuse juillet 2006[PDF], Fédération des chasseurs 55.
- Le cerf élaphe, animaux.org.
- (en) Suttie, J.M., « Social dominance in farmed red deer stags », Applied Animal Behaviour Science, vol. 14, no 2, , p. 191-199 (DOI 10.1016/0168-1591(85)90029-2, lire en ligne)
- (en) Strzeżek, J., Krzywinski, A. et Świdowicz, K., « Seasonal changes in the chemical composition of red deer (Cervus elaphus) semen », Animal Reproduction Science, vol. 9, no 3, , p. 195-204 (DOI 10.1016/0378-4320(85)90002-8, lire en ligne)
- (en) Asher, G.W., Berg, D.K. et Evans, G., « Storage of semen and artificial insemination in deer », Animal Reproduction Science, vol. 62, nos 1-3, , p. 195-211 (DOI 10.1016/S0378-4320(00)00159-7, lire en ligne).
- (en) G. A. Lincoln, Fiona Guinness et R. V. Short, The way in which testosterone controls the social and sexual behavior of the red deer stag (Cervus elaphus), Revue : « Hormones and Behavior » ; vol. 3, no 4, , p. 375-396, DOI:10.1016/0018-506X(72)90027-X (résumé).
- (en) G.W. Asher, J.A. Archer, J.F. Ward, I.C. Scott, R.P. Littlejohn, Effect of melatonin implants on the incidence and timing of puberty in female red deer (Cervus elaphus) ; Animal Reproduction Science, vol. 123, no 3-4, , p. 202-209 ; DOI:10.1016/j.anireprosci.2010.11.018 (Résumé).
- (en) Asher, Geoffrey William, « Reproductive Cycles in Female Cervids », Current Therapy in Large Animal. Theriogenology, , p. 921-931 (DOI 10.1016/B978-072169323-1.50128-8, lire en ligne)
- (en) G.W. Asher, I.C. Scott, J.A. Archer, J.F. Ward et R.P. Littlejohn, Seasonal luteal cyclicity of pubertal and adult red deer (Cervus elaphus) ; Animal Reproduction Science, vol. 125, no 1-4, , p. 138-147, DOI:10.1016/j.anireprosci.2011.03.009.
- (en) G. W. Asher , Effect of subcutaneous melatonin implants on the seasonal attainment of puberty in female red deer (Cervus elaphus) ; Animal Reproduction Science, vol. 22, no 2, , p. 145-159, DOI:10.1016/0378-4320(90)90073-O (Résumé).
- (en) Glenn R. Iason et Fiona E. Guinness, Synchrony of oestrus and conception in red deer (Cervus elaphus L.) ; Animal Behaviour, vol. 33, no 4, , p. 1169-1174, DOI:10.1016/S0003-3472(85)80177-9 (Lien via Science direct).
- (en) Jan M. Bakke et Erik Figenschou, Volatile compounds from the red deer (Cervus elaphus). substances secreted via the urin ; Comparative Biochemistry and Physiology Part A: Physiology, vol. 97, no 3, 1990, p. 427-431, DOI:10.1016/0300-9629(90)90635-6 (Résumé).
- (en) Rolf Langvatn, Prostaglandin (PGF2α-induced parturition in red deer (Cervus elaphus L.) ; Comparative Biochemistry and Physiology Part C: Comparative Pharmacology, vol. 83, no 1, 1986, p. 19-22, (Résumé).
- (en) G.W. Asher, R.C. Mulley, K.T. O’Neill, I.C. Scott, N.B. Jopson et R.P. Littlejohn, Influence of level of nutrition during late pregnancy on reproductive productivity of red deer: I. Adult and primiparous hinds gestating red deer calves. ; Animal Reproduction Science vol. 86, no 3-4, , p. 261-283, DOI:10.1016/j.anireprosci.2004.07.012 (Résumé).
- (en) I.C. Scott, G.W. Asher, J.A. Archer et R.P. Littlejohn, The effect of conception date on gestation length of red deer (Cervus elaphusnext term). ; Animal Reproduction Science, vol. 109, no 1-4, , p. 206-217, DOI:10.1016/j.anireprosci.2007.11.025 (résumé).
- (en) J.F. Aller, O. Fernandez et E. Sanchez, Fixed-time artificial insemination in red deer (Cervus elaphus) in Argentina ; Animal Reproduction Science Volume 115, Issues 1-4, octobre 2009, Pages 312-316 DOI:10.1016/j.anireprosci.2008.11.018 (résumé).
- (en) D. R. J. Bainbridge, S. L. Catt, G. Evans et H. N. Jabbour, Successful in vitro fertilization of in vivo matured oocytes aspirated laparoscopically from red deer hinds ; Theriogenology Volume 51, Issue 5, , pages 891-898 DOI:10.1016/S0093-691X(99)00036-9 (Résumé).
- (en) P.F. Fennessy, G.W. Asher, N.S. Beatson, T.E. Dixon, J.W. Hunter, M.J. Bringans, Embryo transfer in deer ; Theriogenology, Volume 41, Issue 1, , Pages 133-138 ; DOI:10.1016/S0093-691X(05)80058-5 (résumé).
- (en) J.P. Soler, N. Mucci, G.G. Kaiser, J. Aller, J.W. Hunter, T.E. Dixon, R.H. Alberio (2007), Multiple ovulation and embryo transfer with fresh, frozen and vitrified red deer (Cervus elaphus) embryos in Argentina ; Animal Reproduction Science, Volume 102, Issues 3-4, , pages 322-327 ; DOI:10.1016/j.anireprosci.2007.03.024 ; DOI:10.1016/j.theriogenology.2003.11.019 ; (Résumé).
- (en) D. Bowers, C. G. Brown, T. A. Strauch, B. S. Gandy, D. A. Neuendorff, R. D. Randel, S. T. Willard, Artificial insemination following observational versus electronic methods of estrus detection in red deer hinds (Cervus elephus), Original Research ArticleTheriogenology, Volume 62, Issues 3-4, , pages 652-663S. (Résumé).
- (en) Tsenk G. Chassovnikarova, G.G. Markov et P. Müller DNA fingerprinting analysis of red deer (Cervus elaphus L., 1758) populations from Bulgaria ; Mammalian Biology - Zeitschrift fur Saugetierkunde, Volume 68, Issue 6, 2003, pages 394-399 DOI:10.1078/1616-5047-00109 (Lien/Science direct).
- (en) G. W. Asher, K. T. O'Neill, I. C. Scott, B. G. Mockett et M. W. Fisher, Genetic influences on reproduction of female red deer (Cervus elaphus): Seasonal luteal cyclicity. ; Animal Reproduction Science, Volume 59, Issues 1-2, , Pages 43-59 DOI:10.1016/S0378-4320(00)00065-8 (Résumé).
- (en) R. Kuehn, W. Schroeder, F. Pirchner and O. Rottmann, Genetic diversity, gene flow and drift in Bavarian red deer populations (Cervus elaphus) , Conservation Genetics, Volume 4, Number 2, 157-166, DOI:10.1023/A:1023394707884 (Résumé).
- Brachygnathe Brachygnathie Anatomie, odontostomatologie, génétique, embryologie, consulté le .
- (en) F. E. Zachos, C. Althoff, Y. v. Steynitz, I. Eckert et G. B. Hartl (2007) Genetic analysis of an isolated red deer (Cervus elaphus) population showing signs of inbreeding depression ; European Journal of Wildlife Research, Volume 53, Number 1, 61-67, DOI:10.1007/s10344-006-0065-z (Résumé).
- (en) Günther B. Hartl, Frank Zachos et Karl Nadlinger, Genetic diversity in European red deer (Cervus elaphus L.): anthropogenic influences on natural populations Diversité génétique du cerf rouge européen : influences anthropogéniques sur les populations naturelles ; Comptes Rendus Biologies Volume 326, Supplement 1, , Pages 37-42 DOI:10.1016/S1631-0691(03)00025-8 ([Résumé]).
- (en) Simon J. Goodman, Nick H. Barton, Graeme Swanson, Kate Abernethy et Josephine Pemberton, Introgression Through Rare Hybridization: A Genetic Study of a Hybrid Zone Between Red and Sika Deer (Genus Cervus) in Argyll, Scotland (Résumé).
- (en) V. P. W. Lowe, A. S. Gardiner, Hybridization between Red deer (Cervus elaphus) and Sika deer (Cervus nippon) with particular reference to stocks in N.W. England ; Journal of Zoology, Volume 177, Issue 4, pages 553–566, , mis en ligne le , DOI:10.1111/j.1469-7998.1975.tb02259.x (Résumé).
- (en) Simon J. Goodman, Hidetoshi B. Tamate, Rebecca Wilson, Junko Nagata, Shirow Tatsuzawa, Graeme M. Swanson, Josephine M. Pemberton, Dale R. McCullough, Bottlenecks, drift and differentiation: the population structure and demographic history of sika deer (Cervus nippon) in the Japanese archipelago ; Molecular Ecology, Volume 10, Issue 6, pages 1357–1370, et mis en ligne le ; DOI:10.1046/j.1365-294X.2001.01277.x (Résumé).
- (en) Allan D. McDevitt, Ceiridwen J. Edwards, Peter O’Toole, Padruig O’Sullivan, Catherine O’Reilly, Ruth F. Carden, Genetic structure of, and hybridisation between, red (Cervus elaphus) and sika (Cervus nippon) deer in Ireland ; Mammalian Biology - Zeitschrift fur Saugetierkunde, Volume 74, Issue 4, , Pages 263-273 (Résumé).
- (en) Helen V. Senn, Josephine M. Pemberton, Variable extent of hybridization between invasive sika (Cervus nippon) and native red deer (C. elaphus) in a small geographical area ; Molecular Ecology Molecular Ecology ; Volume 18, Issue 5, pages 862–876, et mis en ligne le ; DOI:10.1111/j.1365-294X.2008.04051.x (Résumé).
- (en) J. A. Wass, J. C. Pollard et R. P. Littlejohn, A comparison of the calving behaviour of farmed adult and yearling red deer (Cervus elaphus) hinds. ; Applied Animal Behaviour Science, Volume 80, Issue 4, , Pages 337-345 DOI:10.1016/S0168-1591(02)00236-8 (Résumé).
- (en) J. A. Wass, J. C. Pollard et R. P. Littlejohn, Observations on the hiding behaviour of farmed red deer (Cervus elaphus) calves ; Applied Animal Behaviour Science, Volume 88, Issues 1-2, , Pages 111-120 DOI:10.1016/j.applanim.2004.02.009 (résumé).
- (en) H. Vergar, T. Landete-Castillejos, A. Garcia, P. Molina et L. Gallego, Concentration of Ca, Mg, K, Na, P and Zn in milk in two subspecies of red deer: Cervus elaphus hispanicus and C. e. scoticus ; Small Ruminant Research, Volume 47, Issue 1, , Pages 77-83 DOI:10.1016/S0921-4488(02)00240-7 (Résumé).
- [ranch-amérique.com/cerf-rouge.php ].
- ROC - association reconnue d'utilité publique - le cerf.
- (en) P. N. Grigor, P. J. Goddard et C. A. Littlewood, The movement of farmed red deer through raceways ; Applied Animal Behaviour Science, Volume 52, Issues 1-2, , Pages 171-178 DOI:10.1016/S0168-1591(96)01140-9 (Résumé).
- (en) Cristina B. Sánchez-Prieto, Juan Carranza, Javier Pérez-González, Susana Alarcos et Concha Mateos, Effects of small barriers on habitat use by red deer: Implications for conservation practices ; Journal for Nature Conservation, Volume 18, Issue 3, , Pages 196-201 ; DOI:10.1016/j.jnc.2009.09.002 (Résumé).
- Antoine Morisot et al. (2016) Déplacements des populations de cerfs du massif jurassien : influences climatiques et humaines[PDF] (programme européen Interreg IV-A franco-suisses), Revue Faune sauvage de l'ONCFS, no 103.
- J. G. Jago, R. G. Harcourt et L. R. Matthews, The effect of road-type and distance transported on behaviour, physiology and carcass quality of farmed red deer (Cervus elaphus), Applied Animal Behaviour Science, Volume 51, Issues 1-2, , Pages 129-141 DOI:10.1016/S0168-1591(96)01094-5 (Résumé).
- Saint-Andrieux C., Klein F., Leduc D., Landry P. & B. Guibert (2004), « La progression du Cerf élaphe en France depuis 1985 ». Revue Faune sauvage (ONCFS) no 264 : 19-24.
- Haÿez F. [2006]. « Quand l’ADN parle… » Chasse et nature, 98(9) : 24-28 (5 p., 7 fig.).
- (en) Fajardoa, I. González, I. López-Calleja, I. Martín, M. Rojas, P.E. Hernández, T. García et Rosario Martín, Identification of meats from red deer (Cervus elaphus), fallow deer (Dama dama), and roe deer (Capreolus capreolus) using polymerase chain reaction targeting specific sequences from the mitochondrial 12S rRNA gene ; Meat Science, vol. 76, no 2, , p. 234-240, DOI:10.1016/j.meatsci.2006.11.004 (Résumé).
- (en)Zachos F.E., Althoff C., Steynitz Y.V., Eckert I., Hartl G.B. (2007) - « Genetic analysis of an isolated red deer (Cervus elaphus) population showing signs of inbreeding depression », European Journal of Wildlife Research, 53 : p. 61-67. (résumé).
- La notion de record est relative, car on ne connaît pas vraiment les taux historiques de grands herbivores ni leurs variations depuis la préhistoire ou même lors des glaciations, et il est difficile d'apprécier l'écopotentialité des forêts pour ces espèces, qui varie notamment selon leur productivité biologique, mais aussi la possibilité pour les animaux de changer de zones quand un stade de « surpâturage » est atteint.
- (en) PICHON B, MOUSSON L, FIGUREAU C, RODHAIN F, PÉREZ-EID C. Density of deer in relation to the prevalence of Borrelia burgdorferi s.l. in Ixodes ricinus nymphs in Rambouillet forest, France. Exp. Appl. Acarol. 1999;23:267-275.
- (en) Linton C. Freeman, Sue C. Freeman et A. Kimball Romney, The implications of social structure for dominance hierarchies in red deer, Cervus elaphus L. Animal Behaviour, Volume 44, Part 2, , Pages 239-245 DOI:10.1016/0003-3472(92)90030-D (Résumé).
- (en) B. V. Hodgetts, J. R. Waas et L. R. Matthews, The effects of visual and auditory disturbance on the behaviour of red deer (Cervus elaphus) at pasture with and without shelter Purchase ; Applied Animal Behaviour Science, Volume 55, Issues 3-4, , Pages 337-351 DOI:10.1016/S0168-1591(97)00040-3 (Résumé).
- (en) J. C. Pollard et R. P. Littlejohn, The effects of pen size on the behaviour of farmed red deer stags confined in yards. ; Applied Animal Behaviour Science, Volume 47, Issues 3-4, , Pages 247-253 DOI:10.1016/0168-1591(95)00621-4 ([Résumé]).
- Bertouille S., Buchet R., Manet B. (2007) - « La gestion du cerf non boisé (bilan de la saison 2006) », Chasse et nature 99 (7) : p. 18-24.
- Tableaux de chasse Cerf-Chevreuil-Sanglier. Saison 2006-2007. Supplément au Faune sauvage no 279, ONCFS, 2008.
- Voir sur roc.asso.fr, consulté le .
- (en) Zamora R., « Effect of browsing by ungulates on sapling growth of Scots pine in a mediterranean environment: Consequences for forest regeneration », Forest Ecology and Management, , p. 33-42.
- D'après l'Association des Amis de l'Aigoual du Bougès et du Lozère (site de l'association).
- Quand les loups font pousser les arbres.
- (en) Francisco Ruiz-Fons, Isabel G. Fernández-de-Mera, Pelayo Acevedo, Ursula Höfle, Joaquín Vicente, José de la Fuente et Christian Gortazár, Ixodid ticks parasitizing Iberian red deer (Cervus elaphus hispanicus) and European wild boar (Sus scrofa) from Spain: Geographical and temporal distribution ; Veterinary Parasitology, vol. 140, no 1-2, , p. 133-142, DOI:10.1016/j.vetpar.2006.03.033 (Résumé).
- (en) B.L. Gauss, J.P. Dubey, D. Vidal, O. Cabezón, F. Ruiz-Fons, J. Vicente, I. Marco, S. Lavin, C. Gortazar et S. Almería, Prevalence of Toxoplasma gondii antibodies in red deer (Cervus elaphus) and other wild ruminants from Spain ; Veterinary Parasitology, vol. 136, no 3-4, , p. 193-200, DOI:10.1016/j.vetpar.2005.11.013 (Résumé).
- (en) J.S. Gray, T.M. Murphy, S.M. Taylor, D.A. Blewett et R. Harrington, Comparative morphological and cross transmission studies with bovine and deer babesias in Ireland ; Preventive Veterinary Medicine, vol. 9, no 3, , p. 185-193, DOI:10.1016/0167-5877(90)90065-P ([Résumé]).
- (en) C.G. Mackintosh, R.G. Clark, B. Thompson, B. Tolentino, J.F.T. Griffin et G.W. de Lisle, Age susceptibility of red deer (Cervus elaphus) to paratuberculosis ; Veterinary Microbiology, vol. 143, no 2-4, , p. 255-261, DOI:10.1016/j.vetmic.2009.11.014 (résumé).
- (en) M. Reglero, Mark A. Taggart, Lidia Monsalve-González et Rafael Mateo, Heavy metal exposure in large game from a lead mining area: Effects on oxidative stress and fatty acid composition in liver ; Environmental Pollution, vol. 157, no 4, , p. 1388-1395, The Behaviour and Effects of Nanoparticles in the Environment DOI:10.1016/j.envpol.2008.11.036 (Résumé).
- (en) Knott, J., Gilbert, J., Green, R.E. & Hoccom, D.G. (2009) Comparison of the lethality of lead and copper bullets in deer control operations to reduce incidental lead poisoning; field trials in England and Scotland. Conservation Evidence 6, 71-78. Kokko, H; Laakso, J; Sutherland, W.J. (2009) In memoriam: Professor Esa Ranta (1953-2008) Annales Zoological Fennica 45.
- (en) Alicia M. Hines, Vanessa O. Ezenwa, Paul Cross et Jared D. Rogerson, Effects of supplemental feeding on gastrointestinal parasite infection in elk (Cervus elaphus): Preliminary observations ; Veterinary Parasitology, vol. 148, no 3-4, , p. 350-355, DOI:10.1016/j.vetpar.2007.07.006 (Résumé).
- (en) Francisco Ceacero, Tomás Landete-Castillejos, Andrés J. García, José A. Estévez, Alberto Martinez, Antonio Calatayud, Enrique Gaspar-López et Laureano Gallego, Free-choice mineral consumption in Iberian red deer (Cervus elaphusnext term hispanicus) response to diet deficiencies ; Livestock Science, vol. 122, no 2-3, , p. 345-348, DOI:10.1016/j.livsci.2008.08.002 (Résumé).
- (en) Grażyna Jarzyńska et Jerzy Falandyszlow, Selenium and 17 other largely essential and toxic metals in muscle and organ meats of Red Deer (Cervus elaphusnext term) — Consequences to human health ; Environment International, Volume 37, Issue 5, , pages 882-888 DOI:10.1016/j.envint.2011.02.017 (Résumé).
- Maja Lazarus, Andreja Prevendar Crnić, Nina Bilandžić et Josip Kusak, « Cadmium, Lead, and Mercury Exposure Assessment Among Croatian Consumers of Free-Living Game », Archives of Industrial Hygiene and Toxicology, vol. 65, no 3, , p. 281–292 (ISSN 0004-1254, DOI 10.2478/10004-1254-65-2014-2527, lire en ligne, consulté le )
- (en) U. Kierdorf, H. Kierdorf, M. Erdelen et Z. Machoy, Mandibular bone fluoride accumulation in wild red deer (Cervus elaphusnext term L.) of known age, Comparative Biochemistry and Physiology Part A: Physiology, Volume 110, Issue 4, , pages 299-302 DOI:10.1016/0300-9629(94)00188-Y (Résumé).
- (en) E.J. McGee, H.J. Synnott, C. O'Keefe et P.A. Colgan, Review Radionuclide uptake by red deer(Cervus elaphus) on mountain grazing ; British Veterinary Journal, Volume 151, Issue 6, novembre-, pages 671-682, DOI:10.1016/S0007-1935(95)80148-0 (Résumé).
- (en) Velvet Antler (velours des bois) ; research summary, consulté le .
- (en) « Alternative Uses for Deer Velvet », The Ensign. In: The New Zealand Game Industry Board Media Statement, .
- Jean-Paul Grossin et Antoine Reille, Anthologie du cerf, Paris, Hatier, 1992.
- Michel Pastoureau, Une histoire symbolique du Moyen Âge, Paris, Seuil, 2004, p. 75.
{kind=link}
{kind=link}
Voir aussi
Article connexe
Bibliographie
- (fr) Bonenfant C (2004), Rôle des contraintes évolutives dépendantes du sexe en biologie des populations : l’exemple du Cerf élaphe (Cervus elaphus L.). Thèse de doctorat de l’Université Claude-Bernard (Lyon-I).
- (en) Bonenfant C, Gaillard JM, Klein F & Aillard D (2004), Variation in harem size of red deer (Cervus elaphus L.) : the effect of adult sex ratio and age-structure. Journal of Zoology, 264, 1-9.
- (en) Bonenfant C, Loé LE, Mysterud A, Langvatn R, Stenseth NC, Gaillard JM & Klein F (2004), Multiple causes of sexual segregation in European Red Deer (Cervus elaphus L.) : enlightenments from varying breeding phenology at high and low latitude ; Proceedings of the Royal Society of London, Series B, 269, 1-10.
- (en) Coltman, D.W., Pilkington, J.G. & Pemberton, J.M. 2003. Fine-scale genetic structure in a free living ungulate population. Molécular Ecology 12(3): 733-742. DOI:10.1046/j.1365-294X.2003,01762.x
- (en) Haigh JC (1995) Exploitation and domestication of deer. Anthrozoos: A Multidisciplinary Journal of The Interactions of People & Animals, 8(4), 206-216 (avec ingentaconnect)
- (en) Hebblewhite, M. & Merrill, E.H. 2009. Trade-offs between predation risk and forage differ between migrant strategies in a migratory ungulate. Ecology 90(12): 3445-3454. DOI:10.1890/08-2090.1
- (en) Leblond, M., Dussault, C. & Ouellet, J.-P. 2010. What drives fine-scale movements of large herbivores? A case study using moose. Ecography 33(6): 1102-1112. DOI:10.1111/j.1600-0587.2009.06104.x
- (en) Mantor M (2012) Introduction: Toward Adaptive Management and Assessment in Large Mammal Conservation. Journal of International Wildlife Law & Policy, 15(2), 115-124.
- (en) Mysterud, A., Meisingset, E.L., Veiberg, V., Langvatn, R., Solberg, E.J., Loe, L.E. & Stenseth, N.C. (2007). Monitoring population size of red deer Cervus elaphus: an evaluation of two types of census data from Norway. Wildlife Biology 13: 285-298.
- (en) Putman R, Langbein J, Green P & Watson P (2011) Identifying threshold densities for wild deer in the UK above which negative impacts may occur. Mammal Review, 41(3), 175-196.
- (en) Singh, N.J., Boerger, L., Dettki, H., Bunnefeld, N. & Ericsson, G. 2012. From migration to nomadism: movement variability in a northern ungulate across its latitudinal range. Ecological Applications 22(7): 2007-2020.
- (en) Sykes N, Carden RF & Harris K (2013) Changes in the size and shape of fallow deer—evidence for the movement and management of a species. International Journal of Osteoarchaeology, 23(1), 55-68.
Œuvres
- Biches et cerf au repos - Rosa Bonheur, huile sur toile, 1867
Liens externes
- (fr) Référence INPN : Cervus elaphus Linnaeus, 1758
- (en) Référence Fauna Europaea : Cervus elaphus
- (fr+en) Référence ITIS : Cervus elaphus Linnaeus, 1758
- (en) Référence Animal Diversity Web : Cervus elaphus
- (en) Référence NCBI : Cervus elaphus (taxons inclus)
- (en) Référence UICN : espèce Cervus elaphus Linnaeus, 1758 (consulté le )
- (en) Référence CITES : espèce Cervus elaphus Linnaeus, 1758 (+ répartition sur Species+) (consulté le )
- (en) Référence Fonds documentaire ARKive : Cervus elaphus .
- (en) Référence GISD : espèce Cervus elaphus L., 1758
 Portail des mammifères
Portail des mammifères