Paléontologie
La paléontologie est la discipline scientifique qui étudie les processus de fossilisation (taphonomie) des êtres vivants disparus ou la corrélation et datation des roches qui les contiennent (biostratigraphie). Située au croisement de la géologie et de la biologie, elle décrit l'évolution du monde vivant, les relations entre eux et leur environnement (paléoécologie, évolution de la biosphère), leur répartition spatiale et leurs migrations (paléobiogéographie), l'extinction des espèces et l'apparition de nouvelles, ainsi que les écosystèmes dans lesquels les organismes anciens ont vécu. Parmi ses objectifs figurent, outre la reconstruction d'êtres vivants ayant vécu dans le passé, l'étude de leur origine, de leurs changements dans le temps (évolution et phylogénie).

| Sous-classe de | |
|---|---|
| Partie de | |
| Pratiqué par |
Paléontologue (d) |
| Histoire |
La science s'est établie au XVIIIe siècle grâce aux travaux de Georges Cuvier sur l'anatomie comparée et s'est développée rapidement au XIXe siècle. Au fur et à mesure que les connaissances augmentaient, la paléontologie s'est subdivisé en paléobiologie, taphonomie et biochronologie. Elle fournit des informations nécessaires à d'autres disciplines (étude de l'évolution des êtres vivants, biostratigraphie, paléogéographie ou paléoclimatologie, entre autres).
La paléontologie permet de comprendre la biodiversité et la répartition des êtres vivants sur Terre (biogéographie) - avant l'intervention humaine -, elle a fourni des preuves indispensables pour la solution de deux des plus grandes controverses scientifiques du siècle dernier, l'évolution des êtres vivants et la dérive des continents, et, en vue de notre avenir, elle offre des outils pour analyser comment les changements climatiques peuvent affecter la biosphère dans son ensemble.
Étymologie

Le mot « paléontologie » se décompose en trois parties provenant du grec ancien :
- paleo, de παλαιός, palaios, « ancien » ;
- ontos, de ὄντος, participe-présent du verbe être : « étant » ;
- logie, de λόγος, logos, « l'étude, le discours ».
Il s'agit donc, littéralement, de la « science étudiant la vie ancienne » et, plus précisément, de la discipline qui étudie les organismes disparus ayant laissé dans les terrains sédimentaires des restes de leur corps ou des traces de leurs activités. Ces restes ou traces sont appelés fossiles.
Ce terme a été créé en 1822 par le zoologiste Henri Ducrotay de Blainville (1777-1850) et diffusé en Europe par le géologue britannique Charles Lyell (1797-1875)[1].
Une science au croisement de la géologie et de la biologie
Située au croisement de la géologie et de la biologie[2], la paléontologie décrit l'évolution du monde vivant, l'extinction et l'apparition de certaines espèces[3], ainsi que les écosystèmes dans lesquels les organismes anciens ont vécu[2].
La paléontologie peut être définie comme la science des fossiles[4]. Elle entretient des liens étroits avec la géologie : la datation de ces restes d'organismes vivants repose souvent sur des informations en matière de stratigraphie et sur l'analyse des sédiments[5]. En retour, la paléontologie apporte une contribution importante à la compréhension de l'histoire de la Terre[6]. Ainsi, l'échelle des temps géologiques est divisée en unités définies par les organismes présents, des événements climatiques, etc. : ères, périodes, époques, étages.
La paléontologie est également liée à la biologie. Les deux disciplines ont en partage l'étude des êtres vivants mais ne travaillent pas sur les mêmes données ; la paléontologie n'a accès au vivant qu'à travers les fossiles - archives biologiques -, tandis que l'objet de la biologie est le vivant immédiat[7]. Un des principaux fondateurs de la paléontologie, Georges Cuvier, était un spécialiste de l'anatomie comparée, une branche de la biologie ; ce qui conduit certains historiens des sciences à dire que la paléontologie est née de la biologie (plutôt que de la géologie)[8]. La biologie est mise à contribution par les paléontologues de façon plus récente tout particulièrement dans le cadre de la phylogénétique moléculaire, qui compare l'ADN et ARN des organismes modernes pour reconstruire les "arbres généalogiques" de leurs "ancêtres" ; elle est également mobilisée pour estimer les dates d'importants développements évolutifs, bien que cette approche soit controversée en raison de doutes sur la fiabilité de « l'horloge moléculaire »[9]. Les deux disciplines sont liées plus généralement du fait que les paléontologues utilisent souvent l'observation des caractères prévalant aujourd'hui pour tirer des conclusions sur les mondes d'hier : c'est le principe de l'actualisme[10].
Principales formes de paléontologie
On distingue trois principales formes de paléontologie :
- la « paléontologie systématique » dont l'objectif premier est l'étude des phylogénies sur la base de l'observation scientifique des fossiles ;
- la « paléontologie générale » dite également « paléontologie fondamentale » dont l'objectif premier est la compréhension des problèmes généraux que la démarche systématique permet de découvrir (relations entre les êtres vivants disparus ou actuels, leurs évolutions, et, à plus large échelle, l'évolution des êtres vivants, des milieux et des climats au cours des temps géologiques) ;
- la « paléohistologie » qui est l'étude fine des tissus fossilisés, avec une vaste palette d'objectifs et d'applications, allant de la reconstitution approfondie (comme par exemple la coloration du plumage d'Archéopteryx) à la paléontologie du développement[11], notamment avec l'appui de la phylogénétique moléculaire et assimilés (exemple : comparaison du collagène de Tyrannosaurus rex avec celui des oiseaux actuels[12]).
Les étapes du travail paléontologique

Le travail paléontologique comporte généralement quatre étapes :
- la prospection et les fouilles sur le terrain : c'est la partie la plus ardue, la plus physique, et administrativement compliquée : après obtention de tous les accords nécessaires, du matériel et des fonds, après le transport sur site, il s'agit de quadriller, mesurer, photographier, cartographier, extraire, préserver, emballer les fossiles, tamiser le sédiment, classer les trouvailles, les conditionner pour leur transport… ;
- l'analyse et l'étude en laboratoire (voire dans un accélérateur de particules comme l'ESRF) des fossiles après déballage des colis ; le conditionnement des trouvailles, les moulages, l'attribution des fonctions (collection d'étude, muséologie, échanges…) ;
- la description et publication scientifique des fossiles et des résultats d'étude, la reconstitution des êtres fossilisés et de leurs milieux d'origine ;
- la diffusion pour le grand public des connaissances ainsi acquises (exposition au public, livres, autres publications, documentaires…).
Relations étroites avec d'autres disciplines

Le travail en paléontologie s'effectue en collaboration avec la recherche en archéologie, lorsque les paléontologues (plus exactement les paléoanthropologues) étudient des fossiles humains. La paléontologie identifie aussi les fossiles d'animaux ou de plantes dans les sites archéologiques (nourriture, animaux d'élevage) et analyse le climat contemporain de l'occupation du site[13].
Pour « faire parler » les fossiles, la paléontologie, science largement pluridisciplinaire, emprunte souvent des techniques à d'autres sciences, dont la chimie, l'écologie, la physique et les mathématiques. Ainsi par exemple les signatures géochimiques des roches aident à dater l'apparition de la vie sur Terre[14], et les analyses des rapports isotopiques du carbone peuvent permettre d'identifier les changements climatiques et d'expliquer des transitions majeures telles que l'extinction Permien-Trias[15]. Des techniques d'ingénierie sont utilisées pour analyser la manière dont les corps d'organismes anciens pouvaient fonctionner, par exemple la vitesse de course de Tyrannosaurus et sa force de morsure[16],[17], ou la mécanique de vol de Microraptor[18]. L'étude des détails internes des fossiles a recours à la technique de la microtomographie à rayon X[19],[20]. La paléontologie, la biologie, l'archéologie et la paléoneurobiologie sont associées à l'occasion de l'étude des moulages endocrâniens (endocastes) d'espèces liées à l'homme afin de clarifier l'évolution du cerveau humain[21].
La paléontologie contribue à l'astrobiologie, la recherche d'une vie possible sur d'autres planètes, en développant des modèles de la manière dont la vie a pu apparaître sur Terre et en fournissant des techniques permettant de détecter les preuves d'existence d'êtres vivants[22].
Subdivisions
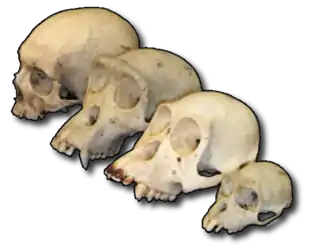
La paléontologie comporte plusieurs sous-disciplines. La paléontologie des vertébrés étudie les fossiles des premiers poissons, jusqu'à ceux des ancêtres immédiats des mammifères modernes. Bien que l'homme fasse partie des vertébrés, son étude est considérée comme un champ distinct, la paléoanthropologie, qui synthétise des connaissances provenant d'autres disciplines comme l'anthropologie et l'archéologie. La paléontologie des invertébrés traite de fossiles tels que ceux des mollusques, des arthropodes, des vers et les échinodermes. La micropaléontologie se focalise sur les microfossiles[23]. Les microfossiles qui ne sont pas minéralisés, mais organiques, tels que les grains de pollen, les spores, font l'objet d'une sous-discipline séparée, la palynologie (ou paléopalynologie). La paléobotanique, étudiant les plantes fossiles, est très proche de la palynologie, mais s'en distingue par les restes botaniques qu'elle prend pour objet - non pas des "poussières" végétales mais le bois pétrifié, la houille (provenant de la carbonisation de végétaux), des impressions de feuilles dans la roche[24]. La paléoichnologie se consacre aux traces laissées par les animaux (pistes, terriers…), la paléocoprologie à l'étude des excréments.
La paléontologie se diversifie, depuis les années 1960, en participant à des approches fondamentalement pluridisciplinaires qui deviennent autant de disciplines nouvelles et interconnectées : la paléoécologie, la paléoclimatologie, la biostratigraphie, la paléobiogéographie, etc.
Au lieu de se concentrer sur les organismes individuels, la paléoécologie examine les interactions entre différents organismes anciens, tels que leurs chaînes alimentaires, et les interactions avec leur environnement[25].
La paléoclimatologie, bien que parfois considérée comme une sous-discipline de la paléoécologie, se concentre davantage sur l'histoire du climat de la Terre et sur les mécanismes qui l'ont modifié[26]. Ces modifications peuvent être liées à des développements évolutifs, comme l'expansion rapide des plantes terrestres durant le Dévonien qui a éliminé une plus grande quantité de dioxyde de carbone de l'atmosphère, réduisant l'effet de serre et contribuant ainsi à provoquer un âge glaciaire durant le Carbonifère[27].
La biostratigraphie, utilisant les fossiles pour déterminer l'ordre chronologique dans lequel les roches se sont formées, est utile aux paléontologues et aux géologues[28].
La paléobiogéographie étudie la distribution spatiale des organismes ; elle est également liée à la géologie. La répartition géographique des fossiles renvoie en effet à des mouvements de migration et permet de montrer des connexions entre des îles ou des continents[29].
La taphonomie est une branche de la paléontologie qui s'intéresse aux processus de fossilisation, qui font intervenir notamment la biodégradation et, après l'enfouissement de l'organisme, les effets de la diagenèse[30].


 Paléobotanique : empreinte de feuille de fougère.
Paléobotanique : empreinte de feuille de fougère. Paléoécologie : Bryozoa et Brachiopoda, Ordovician.
Paléoécologie : Bryozoa et Brachiopoda, Ordovician. Taphonomie : Perforations d'éponges et tubes encroutés de vers serpulidae sur une coquille moderne de Mercenaria de Caroline du Nord.
Taphonomie : Perforations d'éponges et tubes encroutés de vers serpulidae sur une coquille moderne de Mercenaria de Caroline du Nord.
Interprétation des fossiles

Les fossiles des organismes anciens constituent généralement le type de preuve le plus décisif en paléontologie. Les fossiles les plus communs sont les os, les coquillages, le bois[31]. Ils ont généralement un caractère fragmentaire : ce sont par exemple des éléments isolés de squelettes, et parfois de simples traces (empreintes, terriers). La fossilisation étant un phénomène rare, et la plupart des organismes fossilisés ayant été détruits au cours du temps par érosion, ou par métamorphisme, les archives fossiles sont par conséquent très incomplètes. Le paléontologue doit prendre en considération les biais dans les archives fossiles : certains environnements différents sont plus favorables que d'autres à la fossilisation ; et les organismes avec un squelette minéralisé ayant plus de chances d'être conservés, sont donc surreprésentés par rapport aux organismes à corps mou[32]. Ainsi, bien qu'il existe plus de 30 phylums d'animaux vivants, les deux tiers n'ont jamais été trouvés sous forme de fossiles[33].
Parfois, des environnements inhabituels, les Lagerstätten, préservent les tissus mous. Cependant, même les Lagerstätten présentent une image incomplète de la vie au cours des temps géologiques, et la majorité des organismes vivants anciens n'y sont probablement pas représentés, parce que les Lagerstätten sont limités à un type particulier d'environnements, ceux où les organismes à corps mou peuvent être préservés très rapidement par des événements exceptionnels comme des glissements de terrain, qui entraînent un enterrement immédiat[34].
Classification des organismes
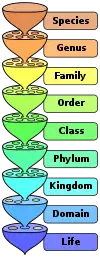
Nommer les groupes d'organismes d'une manière claire est important ; les malentendus sur les noms peuvent conduire à des divergences considérables dans les interprétations scientifiques[35]. La taxonomie linnéenne, couramment utilisée pour classer les organismes vivants, se heurte à des difficultés lorsqu'il s'agit de la classification d'organismes anciens significativement différents des organismes connus. Ainsi, il est parfois difficile de savoir à quel niveau placer un groupe nouvellement découvert, et de dire si ce groupe constitue un genre, une famille ou un ordre ; dans ce mode de classification classique, lorsqu'un groupe est déplacé vers un niveau différent, il doit être renommé[36].
| Tétrapodes |
| ||||||||||||||||||||||||||||||||||||||||||
(bg) Le « sang chaud » est un caractère apparu à un moment de la transition
synapsides–mammifères.
? Le « sang chaud » a dû également apparaître à un de ces moments de
l'évolution – un exemple de convergence évolutive[37].
Les paléontologues utilisent généralement des approches fondées sur la cladistique, une méthode permettant d'élaborer « l'arbre généalogique » évolutif d'un ensemble d'organismes[35]. Selon la théorie cladistique, si les groupes B et C ont plus de similitudes entre eux que l'un ou l'autre n'en a avec le groupe A, alors B et C sont plus étroitement liés l'un à l'autre que l'un ou l'autre avec A. Les caractères qui sont comparés peuvent être anatomiques (comme, par exemple, la présence d'une chorde), ou moléculaires (en se fondant notamment sur la confrontation de séquences d'ADN ou de protéines). Le résultat d'une analyse réussie est une hiérarchie de clades — des groupes qui partagent un ancêtre commun. La technique cladistique est faillible, car certaines caractéristiques, telles que les ailes, ont évolué plus d'une fois ; l'analyse doit prendre en compte la possibilité d'apparitions multiples de certains caractères de manière indépendante dans différentes lignées, phénomène appelé convergence évolutive[37].
L'un des objectifs de la paléontologie est de reconstituer l'histoire de l'évolution. Cependant, le but n'est pas de découvrir les espèces intermédiaires entre deux espèces car il n'y a pas d'intermédiaires-ancêtres à trouver mais des intermédiaires structuraux à définir. Les Archaeopteryx et les oiseaux de l'ère secondaire, tout comme les poissons à poumons par exemple, sont des intermédiaires structuraux. Les probabilités de trouver « les ancêtres réels » des chaînes généalogiques sont infimes en paléontologie, on se « contente » des intermédiaires structuraux pour confirmer ou infirmer les « modèles généalogiques ».
Quelques exemples d'histoires évolutives :
Datation des organismes
velatus
hippocrepis
tiziani
subbullatus
americanus
multibrachiatus
americanus
niagarense
planicosta
subcircularis
bosei
proliferum
unicornis
La paléontologie cherche à retracer l'évolution des êtres vivants, ce qui suppose de les situer chronologiquement. Les paléontologues peuvent prendre appui, pour dater les fossiles, sur la stratigraphie, science du déchiffrement de la succession des couches sédimentaires[38]. Les roches forment habituellement des couches horizontales, chaque couche étant plus récente que celle qu'elle recouvre. Si un fossile apparaît entre deux couches dont les âges sont connus, l'âge du fossile doit se situer entre ces deux âges connus[39]. Cependant, les séquences de roches ne sont pas continues, et peuvent être interrompues par des failles ou des périodes d'érosion. Des fossiles d'espèces ayant vécu pendant un intervalle relativement court peuvent alors être utilisés pour corréler des roches isolées : cette technique est appelée biostratigraphie. Par exemple, le conodonte Eoplacognathus pseudoplanus a vécu dans la période de l'Ordovicien moyen[40]. Si des roches d'âge inconnu présentent des restes d' E. Pseudoplanus, on en déduit qu'elles doivent dater de l'Ordovicien moyen. Un bon fossile stratigraphique doit, pour être utile, être réparti dans plusieurs régions du monde et appartenir à une espèce ayant vécu durant une courte période. La stratigraphie et la biostratigraphie ne peuvent fournir qu'une datation relative (A avant B), ce qui est souvent suffisant pour étudier l'évolution.
Les lits qui conservent des fossiles manquent généralement des éléments radioactifs nécessaires à la datation radiométrique. Cette technique est le seul moyen de déterminer l'âge absolu des roches vieilles de plus de 50 millions d'années ; elle peut être précise à 0,5% ou même davantage[41]. Le principe de la datation radiométrique est simple : les vitesses de désintégration de divers éléments radioactifs sont connus, et ainsi le rapport de l'élément radioactif à l'élément dans lequel il se désintègre montre depuis combien de temps l'élément radioactif a été incorporé dans la roche. Les éléments radioactifs ne sont communs que dans les roches d'origine volcanique ; les seules roches renfermant des fossiles qui peuvent être datées par radiométrie sont donc les couches de cendres volcaniques[41].
Les relations dans l'arbre généalogique peuvent également aider à déterminer la date d'apparition des lignées. Il est possible d'estimer depuis combien de temps deux clades vivants ont divergé - c'est-à-dire approximativement depuis combien de temps leur dernier ancêtre commun a dû vivre - en supposant que les mutations d'ADN s'accumulent à un rythme constant. Ces « horloges moléculaires » sont cependant faillibles et ne fournissent qu'un « timing » très approximatif[42].
Histoire évolutive du vivant
La paléontologie retrace l'histoire évolutive du vivant, qui date de 3,4 milliards d'années, peut-être de 3,7 milliards d'années, voire davantage selon certains scientifiques — la Terre, elle, ayant été formée il y a 4,5 milliards d'années. Les fossiles les plus anciens sont souvent controversés, et leur origine biologique mise en cause, mais les milieux scientifiques admettent assez généralement que des stromatolithes australiens, datant de 3,4 milliards d'années, résultant de l'activité métabolique de colonies de bactéries, sont la preuve de vie la plus ancienne. Selon la théorie de la panspermie, la vie sur Terre aurait été "ensemencée" par des météorites[43], mais la plupart des chercheurs actuels pensent que la vie a surgi sur Terre et non par un transfert d'organismes vivants à travers l'espace[44].
Pendant environ deux milliards d'années, des tapis microbiens, colonies multicouches de différentes bactéries, ont constitué la forme dominante de la vie sur Terre[45]. L'évolution de la photosynthèse oxygénée leur a permis de jouer un rôle majeur dans l'oxygénation de l'atmosphère[46] depuis environ 2,4 milliards d'années. Ce changement d'atmosphère a augmenté leur efficacité en tant que pépinières de l'évolution[47]. Si les eucaryotes, cellules avec des structures internes complexes, peuvent avoir été présents plus tôt, leur évolution s'est accélérée quand ils ont acquis la capacité de transformer l'oxygène et d'en faire une source puissante d'énergie métabolique. Cette innovation peut provenir d'eucaryotes primitifs capturant des bactéries alimentées par l'oxygène et les transformant en organites appelées mitochondries[48]. La première preuve de l'existence d'eucaryotes complexes avec des organites (comme les mitochondries) date d'il y a 1,85 milliard d'années[49],[50].
Les premiers fossiles d'organismes multicellulaires (composés de cellules eucaryotes), sont les Gabonionta, datant de 2,1 milliards d'années. La spécialisation des cellules dans l'accomplissement de différentes fonctions apparaît pour la première fois il y a 1,43 milliard d'années avec un possible champignon, et il y a 1,2 milliard d'années, avec probablement une algue rouge.
Les premiers animaux connus sont des cnidaires d'il y a environ 580 millions d'années, mais selon les scientifiques, ils doivent descendre d'animaux antérieurs[51]. Les premiers fossiles d'animaux sont rares parce qu'ils n'avaient pas développé de parties dures minéralisées et facilement fossilisées jusqu'à il y a environ 548 millions d'années[52]. Les premiers animaux bilatériens d'apparence moderne apparaissent au Cambrien inférieur. Il y a un débat de longue date pour savoir si l'explosion cambrienne était vraiment une période très rapide d'expérimentation évolutionnaire ; selon d'autres hypothèses les animaux d'apparence moderne auraient commencé à évoluer plus tôt mais les fossiles de leurs précurseurs n'ont pas encore été découverts[53]. Les vertébrés sont restés un groupe mineur jusqu'à l'Ordovicien supérieur[54],[55].
Paléontologues célèbres
- Georges Cuvier (1769-1832)
- Philippe-Charles Schmerling (1791-1836)
- Alcide Dessalines d'Orbigny (1802-1857)
- Louis Édouard Gourdan de Fromentel (1824-1901)
- Henry Testot-Ferry (1826-1869)
- Othniel Charles Marsh (1831-1899)
- Edward Drinker Cope (1840-1897)
- Franz Nopcsa (1877-1933)
- Pierre Teilhard de Chardin (1881-1955)
- Paul Wernert (1889-1972)
- Véra Gromova (1891-1973)
- Louis Seymour Bazett Leakey (1903-1972)
- René Lavocat (1909-2007)
- Jean-Pierre Lehman (1914-1981)
- Sándor Bökönyi (1926-1994) paléozoologue
- Yves Coppens (1934- 2022)
- Henry de Lumley (1934-)
- Marie-Françoise Bonifay (1935-)
- Philippe Taquet (1940-)
- Michel Brunet (1940-)
- Stephen Jay Gould (1941-2002)
- Robert T. Bakker (1945-)
- Jack Horner (1946-)
- Éric Buffetaut (1950-)
- Brigitte Senut (1954-)
- Neil Shubin (1960-)
- Francis Duranthon (1961-)
- Ross Geller (1966-)
- José Braga (1967-)
La reconstitution paléontologique

Les paléontologues reconstituent, à partir des fossiles et par les méthodes de l'anatomie comparée inaugurée par Georges Cuvier, la morphologie des plantes et des animaux disparus. La palynologie leur permet de reconstituer la flore, l'ichnologie - les déplacements, les dentitions et les coprolithes - les régimes alimentaires, divers autres indices - les climats... mais une fois cela établi, des artisans, illustrateurs, ingénieurs du son ou cinéastes doivent « mettre en vie » les reconstitutions. Leur rôle est très important, leurs réalisations ont marqué les imaginations, suscité des vocations, permis le financement d'expéditions. Dès le XIXe siècle, à Londres, les reconstitutions du Crystal Palace ont créé un engouement pour les Dinosaures, et les vues d'artiste d'espèces préhistoriques, polychromes ou non, se dressent souvent à l'entrée des musées à vocation paléontologique (comme devant la galerie de Paléontologie et d'Anatomie comparée du Muséum national d'histoire naturelle, à Paris) ou en illustrent l'intérieur (comme au Musée d'histoire naturelle de Londres)[56].
Généralement, les fossiles ne donnent pas d'indication des couleurs, et celles-ci ne peuvent être qu'imaginées à l'exemple des êtres vivants actuels occupant des milieux homologues et ayant une éthologie comparable. Il en est de même pour les sons : si, exceptionnellement, certains crânes d'hadrosaures dans les crêtes desquels on a insufflé de l'air, ont pu produire une gamme de sonorités, le rythme et l'harmonie en resteront à jamais inconnus : là encore, on les imagine d'après les sons produits par les animaux actuels apparentés (on devrait d'ailleurs représenter les dinosaures plutôt sifflant comme des oiseaux, que rugissant comme des lions ou meuglant comme des bovins). Les reconstitutions (ou « reconstructions ») paléontologiques sont donc par définition, et l'on doit en avertir les publics, en partie conjecturelles, puisque même dans les cas rares de conservation dans l'ambre ou la glace, l'organisme est dégradé notamment au niveau chromatique ; il s'agit par la force des faits, de vues d'artiste, mais des vues qui, pour rester dans les limites de ce qui est admis par les chercheurs au moment où elles sont réalisées, doivent obligatoirement s'inspirer d'autres images déjà scientifiquement validées dans les publications spécialisées. Si l'on ne comprend pas ce processus de « conjecture restitutive », on ne peut que considérer comme « inexactes » ou « violant quelque copyright » la totalité des vues d'artiste, d'images de synthèse, de reconstitutions en volume existantes[57], et l'on amputera la paléontologie de toute son iconographie « grand public » y compris les œuvres de maîtres comme Mauricio Antón, Alain Bénéteau, Dimitri Bogdanov, Zdeněk Burian, Heinrich Harder, Charles R. Knight ou Édouard Riou, seules échappant à ce « grand ménage » les illustrations strictement scientifiques des articles spécialisés.
Tant qu'illustrateurs, artisans ou réalisateurs de films se conforment aux indications des paléontologues, la reconstitution paléontologique est une simple représentation documentaire, cadrée par la démarche scientifique et qui ne vise qu'à une représentation la plus réaliste possible et limite la créativité personnelle. Mais cette représentation peut sortir du cadre documentaire et entrer alors dans le domaine de l'art, le plus souvent dans la science-fiction (tels le livre et le film Jurassic Park) ou dans l'« heroic fantasy » (tels le livre et la série Dinotopia).
Dans la culture
Dans leurs conférences et interviews, Guillaume Lecointre et Pascal Tassy soulignent que beaucoup de mythes anciens ont pu apparaître à la suite d'observations paléontologiques ou autres, mais sans analyse scientifique[58] :
- des fossiles marins en montagne ont pu donner lieu aux récits de déluges ;
- des ammonites Ceratitida à enroulement incomplet ont été comprises comme des cornes de béliers géants (d'où leur nom) ;
- des ossements de grande dimension ont accrédité l’existence de dragons, de trolls, de titans... ;
- des crânes d’Elephas falconeri ont pu être interprétés comme des têtes de cyclopes (la fosse nasale étant prise pour une orbite unique)[59],[60] et notamment par le philosophe, poète, ingénieur et médecin grec de Sicile, du Ve siècle av. J.-C. Empédocle[61] ;
- des crânes de gigantopithèques (perdus pour la science, qui ne dispose que de dents) ont pu être à l'origine du mythe du yéti ;
- les volcans et les laves, comme bouches des enfers ;
- des tentacules flottants d’Architeuthis (décrit scientifiquement en 1857 par Japetus Steenstrup), pouvaient être interprétés comme des serpents de mer géants ou des restes de kraken ;
- des tourbillons associés à des céphalopodes géants ont pu donner le mythe de Charybde avalant l'eau et la recrachant sur les navires et de Scylla aux nombreux bras qui saisissent les marins[62] ;
- les éclairs et la foudre ont pu être interprétés comme des colères divines.
En ce sens l’étude de la mythologie en relation avec la paléontologie, peut parfois mener les chercheurs à de nouvelles découvertes de fossiles, parfois même déjà en collection dans un établissement scientifique, comme dans le cas de Léonard Ginsburg qui retrouve en 1984, dans des collections anciennes, une caisse contenant des fossiles de grande dimension, qu'un forain du XVIIIe siècle avait présenté à travers la France comme étant « les ossemens du géant Theutobocus, roy des Teutons, tué par Marius à la bataille d'Aix en Provence » : il en identifie une dent comme étant celle d'un Dinothère, un éléphant disparu. Le mythe du géant Theutobocus, diffusé au XVIe siècle par Mazuyer, chirurgien à Beaurepaire, et par David Bertrand ou Chenevier, notaire, avait déjà été dénoncé au XVIIe siècle par un autre chirurgien, Jean Riolan, et au XIXe siècle par l'anatomiste Blainville[63].
Plus récemment, les paléontologues ont souvent été popularisés comme « chasseurs de fossiles » : une pratique d'amateurs qui est parfois pratiquée illégalement, éventuellement au détriment des archives fossiles et données scientifiques (déterrage sauvage, dégradation du contexte fossilisé, non-déclaration des « trouvailles », vol sur sites de fouille, trafic, etc), et qui doit beaucoup à la médiatisation des expéditions de Cope et Marsh aux États-Unis. L'œuvre cinématographique Jurassic Park a aussi contribué à populariser l'existence de la paléohistologie dans les fictions modernes, concurrençant l'autre principale source de fantasmes à « fossiles vivants » que sont les « mondes perdus » et la « résurrection » d'espèces éteintes.
Notes et références
- Alain Rey, Dictionnaire historique de la langue française, Le Robert,
- Thierry Tortosa, Principes de paléontologie, Dunod, (lire en ligne), p. 1 ; 6
- Édouard Boureau, Patrick De Wever, Jean Piveteau, « Paléontologie », Encyclopædia Universalis, consulté le 28 juin 2020.
- "La paléontologie a pour objet les fossiles", Thierry Tortosa, Principes de paléontologie, Dunod, (lire en ligne), p. 4
- « Paléontologie », sur futura-sciences.com (consulté le )
- Thierry Tortosa, Principes de paléontologie, Dunod, (lire en ligne), p. 6
- Léna Alex, « L’archive biologique en question : le paléontologue est-il un historien ou un biologiste ? », Bulletin d’histoire et d’épistémologie des sciences de la vie, 2013/2 (Volume 20), p. 197-213. DOI : 10.3917/bhesv.202.0197. URL : https://www.cairn.info/revue-bulletin-d-histoire-et-d-epistemologie-des-sciences-de-la-vie-2013-2-page-197.htm
- Goulven Laurent, « Le développement de la paléontologie », sur futura-sciences.com (consulté le )
- (en) Peterson, Kevin J. et Butterfield, N.J., « Origin of the Eumetazoa: Testing ecological predictions of molecular clocks against the Proterozoic fossil record », Proceedings of the National Academy of Sciences, vol. 102, no 27, , p. 9547–52 (PMID 15983372, PMCID 1172262, DOI 10.1073/pnas.0503660102, Bibcode 2005PNAS..102.9547P)
- Thierry Tortosa, Principes de paléontologie, Dunod, (lire en ligne), p. 5
- (en) « "developmental-palaeontology" »
- « Le collagène de T-Rex nous désigne ses cousins », sur futura-sciences.com
- (en) « How does paleontology differ from anthropology and archaeology? » [archive du ], University of California Museum of Paleontology (consulté le )
- (en) Brasier, M., McLoughlin, N., Green, O. et Wacey, D., « A fresh look at the fossil evidence for early Archaean cellular life », Philosophical Transactions of the Royal Society B, vol. 361, no 1470, , p. 887–902 (PMID 16754605, PMCID 1578727, DOI 10.1098/rstb.2006.1835, lire en ligne [archive du ], consulté le )
- (en) Twitchett RJ, Looy CV, Morante R, Visscher H et Wignall PB, « Rapid and synchronous collapse of marine and terrestrial ecosystems during the end-Permian biotic crisis », Geology, vol. 29, no 4, , p. 351–354 (DOI 10.1130/0091-7613(2001)029<0351:RASCOM>2.0.CO;2, Bibcode 2001Geo....29..351T, lire en ligne)
- (en) Hutchinson, J. R. et Garcia, M., « Tyrannosaurus was not a fast runner », Nature, vol. 415, no 6875, , p. 1018–1021 (PMID 11875567, DOI 10.1038/4151018a, Bibcode 2002Natur.415.1018H) Summary in press release No Olympian: Analysis hints T. rex ran slowly, if at all « https://web.archive.org/web/20080415183735/http://www.sciencenews.org/articles/20020302/fob1.asp »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?),
- (en) M.B. Meers, « Maximum bite force and prey size of Tyrannosaurus rex and their relationships to the inference of feeding behavior », Historical Biology, vol. 16, no 1, , p. 1–12 (DOI 10.1080/0891296021000050755)
- (en) « The Four Winged Dinosaur: Wind Tunnel Test », NOVA (consulté le )
- (en) Russell J. Garwood, Imran A. Rahman et Mark D. A. Sutton, « From clergymen to computers: the advent of virtual palaeontology », Geology Today, vol. 26, no 3, , p. 96–100 (DOI 10.1111/j.1365-2451.2010.00753.x, lire en ligne, consulté le )
- (en) Mark Sutton, Imran Rahman et Russell Garwood, Techniques for Virtual Palaeontology, Wiley, (ISBN 978-1-118-59125-3, lire en ligne)
- (en) Emiliano Bruner, « Geometric morphometrics and palaeoneurology: brain shape evolution in the genus Homo », Journal of Human Evolution, vol. 47, no 5, , p. 279–303 (PMID 15530349, DOI 10.1016/j.jhevol.2004.03.009)
- (en) Cady, S.L., « Astrobiology: A New Frontier for 21st Century Paleontologists », PALAIOS, vol. 13, no 2, , p. 95–97 (PMID 11542813, DOI 10.2307/3515482, JSTOR 3515482, Bibcode 1998Palai..13...95C)
- (en) « What is Paleontology? » [archive du ], University of California Museum of Paleontology (consulté le )
- Thierry Tortosa, Principes de paléontologie, Dunod, (lire en ligne), p. 8
- (en) Kitchell, J.A., « Evolutionary Paleocology: Recent Contributions to Evolutionary Theory », Paleobiology, vol. 11, no 1, , p. 91–104 (DOI 10.1017/S0094837300011428, lire en ligne [archive du ], consulté le )
- (en) « Paleoclimatology » [archive du ], Ohio State University (consulté le )
- (en) Algeo, T.J. et Scheckler, S.E., « Terrestrial-marine teleconnections in the Devonian: links between the evolution of land plants, weathering processes, and marine anoxic events », Philosophical Transactions of the Royal Society B, vol. 353, no 1365, , p. 113–130 (PMCID 1692181, DOI 10.1098/rstb.1998.0195)
- (en) « Biostratigraphy: William Smith » [archive du ] (consulté le )
- Thierry Tortosa, Principes de paléontologie, Dunod, (lire en ligne), p. 9-10
- Thierry Tortosa, Principes de paléontologie, Dunod, (lire en ligne), p. 10, 12
- (en) « What is paleontology? » [archive du ], University of California Museum of Paleontology (consulté le )
- Thierry Tortosa, Principes de paléontologie, Dunod, (lire en ligne), p. 14
- Cowen, R., History of Life, Blackwell Science, , 3rd éd. (ISBN 0-632-04444-6), p. 61
- (en) Butterfield, N.J., « Ecology and evolution of Cambrian plankton »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?), Columbia University Press, New York, (consulté le ), p. 200–216
- (en) Brochu, C.A et Sumrall, C.D., « Phylogenetic Nomenclature and Paleontology », Journal of Paleontology, vol. 75, no 4, , p. 754–757 (ISSN 0022-3360, DOI 10.1666/0022-3360(2001)075<0754:PNAP>2.0.CO;2, JSTOR 1306999)
- (en) Ereshefsky, M., The Poverty of the Linnaean Hierarchy: A Philosophical Study of Biological Taxonomy, Cambridge University Press, (ISBN 0-521-78170-1, lire en ligne), p. 5
- (en) Cowen, R., History of Life, Blackwell Science, , 3rd éd., 47–50 p. (ISBN 0-632-04444-6)
- (en) Pufahl, P.K., Grimm, K.A., Abed, A.M. et Sadaqah, R.M.Y., « Upper Cretaceous (Campanian) phosphorites in Jordan: implications for the formation of a south Tethyan phosphorite giant », Sedimentary Geology, vol. 161, nos 3–4, , p. 175–205 (DOI 10.1016/S0037-0738(03)00070-8, Bibcode 2003SedG..161..175P)
- (en) « Geologic Time: Radiometric Time Scale » [archive du ], U.S. Geological Survey (consulté le )
- (en) Löfgren, A., « The conodont fauna in the Middle Ordovician Eoplacognathus pseudoplanus Zone of Baltoscandia », Geological Magazine, vol. 141, no 4, , p. 505–524 (DOI 10.1017/S0016756804009227, Bibcode 2004GeoM..141..505L)
- (en) Martin, M.W., Grazhdankin, D.V., Bowring, S.A., Evans, D.A.D., Fedonkin, M.A. et Kirschvink, J.L., « Age of Neoproterozoic Bilaterian Body and Trace Fossils, White Sea, Russia: Implications for Metazoan Evolution » (abstract), Science, vol. 288, no 5467, , p. 841–5 (PMID 10797002, DOI 10.1126/science.288.5467.841, Bibcode 2000Sci...288..841M, lire en ligne)
- (en) Hug, L.A. et Roger, A.J., « The Impact of Fossils and Taxon Sampling on Ancient Molecular Dating Analyses », Molecular Biology and Evolution, vol. 24, no 8, , p. 889–1897 (PMID 17556757, DOI 10.1093/molbev/msm115)
-
- (en) Arrhenius, S., « The Propagation of Life in Space », Die Umschau, vol. 7, , p. 32 (Bibcode 1980qel..book...32A) Reprinted in The Quest for Extraterrestrial Life, University Science Books (ISBN 0-19-855704-3)
* (en) Hoyle, F. et Wickramasinghe, C., « On the Nature of Interstellar Grains », Astrophysics and Space Science, vol. 66, no 1, , p. 77–90 (DOI 10.1007/BF00648361, Bibcode 1979Ap&SS..66...77H)
* (en) F. H. Crick et L. E. Orgel, « Directed Panspermia », Icarus, vol. 19, no 3, , p. 341–348 (DOI 10.1016/0019-1035(73)90110-3, Bibcode 1973Icar...19..341C)
- (en) Arrhenius, S., « The Propagation of Life in Space », Die Umschau, vol. 7, , p. 32 (Bibcode 1980qel..book...32A) Reprinted in The Quest for Extraterrestrial Life, University Science Books (ISBN 0-19-855704-3)
- (en) Peretó, J., « Controversies on the origin of life », Int. Microbiol., vol. 8, no 1, , p. 23–31 (PMID 15906258, lire en ligne [archive du ], consulté le )
- (en) Krumbein, W.E., Brehm, U., Gerdes, G., Gorbushina, A.A., Levit, G. et Palinska, K.A., Fossil and Recent Biofilms: A Natural History of Life on Earth, Kluwer Academic, , 1–28 p. (ISBN 1-4020-1597-6, lire en ligne [archive du ])
- (en) Hoehler, T.M., Bebout, B.M. et Des Marais, D.J., « The role of microbial mats in the production of reduced gases on the early Earth », Nature, vol. 412, no 6844, , p. 324–327 (PMID 11460161, DOI 10.1038/35085554, Bibcode 2001Natur.412..324H)
- (en) Nisbet, E.G. et Fowler, C.M.R., « Archaean metabolic evolution of microbial mats », Proceedings of the Royal Society B, vol. 266, no 1436, , p. 2375 (PMCID 1690475, DOI 10.1098/rspb.1999.0934)
- (en) Gray MW, Burger G et Lang BF, « Mitochondrial evolution », Science, vol. 283, no 5407, , p. 1476–81 (PMID 10066161, DOI 10.1126/science.283.5407.1476, Bibcode 1999Sci...283.1476G)
- (en) Andrew H. Knoll, Emmanuelle J. Javaux, David Hewitt et Phoebe Cohen, « Eukaryotic organisms in Proterozoic oceans », Philosophical Transactions of the Royal Society B, vol. 361, no 1470, , p. 1023–1038 (ISSN 0962-8436, PMID 16754612, PMCID 1578724, DOI 10.1098/rstb.2006.1843)
- (en) Mikhail A. Fedonkin, « The origin of the Metazoa in the light of the Proterozoic fossil record », Paleontological Research, vol. 7, no 1, , p. 9–41 (ISSN 1342-8144, DOI 10.2517/prpsj.7.9, S2CID 55178329, lire en ligne [archive du ])
- (en) Chen, J.-Y., Oliveri, P., Gao, F., Dornbos, S.Q., Li, C-W., Bottjer, D.J. et Davidson, E.H., « Precambrian Animal Life: Probable Developmental and Adult Cnidarian Forms from Southwest China », Developmental Biology, vol. 248, no 1, , p. 182–196 (PMID 12142030, DOI 10.1006/dbio.2002.0714, lire en ligne [archive du ], consulté le )
- (en) Bengtson, S. et Waggoner, B.M., « Early Skeletal Fossils », The Paleontological Society Papers, Lipps, J.H., vol. 10 Neoproterozoic—Cambrian Biological Revolutions, , p. 67–78 (DOI 10.1017/S1089332600002345, lire en ligne [archive du ], consulté le )
- (en) Marshall, C.R., « Explaining the Cambrian "Explosion" of Animals », Annu. Rev. Earth Planet. Sci., vol. 34, , p. 355–384 (DOI 10.1146/annurev.earth.33.031504.103001, Bibcode 2006AREPS..34..355M, lire en ligne)
- (en) Conway Morris, S., « Once we were worms », New Scientist, vol. 179, no 2406, , p. 34 (lire en ligne [archive du ], consulté le )
- (en) Sansom I.J., Smith, M.M. et Smith, M.P., Major Events in Early Vertebrate Evolution, Taylor and Francis, , 156–171 p. (ISBN 0-415-23370-4)
- Ion Galtier et Stephen Giner (préf. Éric Buffetaut), Miroirs de la Terre histoire géonomique de la Provence et du Var, notre pays au fil des ères et des climats, Toulon, Les Presses du Midi, , 282 p. (ISBN 978-2-812-70188-7, OCLC 758915939), p. 14, 15, 129-133
- C'est le cas d'un grand nombre d'images paléontologiques, mais aussi archéologiques ou historiques, téléchargées sur Wikimedia Commons.
- Conférence gratuite « Paléontologie, mythes et légendes », à l'auditorium de la grande galerie de l'Évolution du Muséum national d'histoire naturelle le 18 juin 2018 par Pascal Tassy : .
- Linda Gamlin, L'Evolution : les transformations du vivant, Paris, Editions Gallimard, coll. « Les Transformations du vivant » (no 6), , 63 p. (ISBN 978-2-070-57986-0, OCLC 1000086128)
- Stéphane Deligeorges, « Des éléphants de 90 cm au garrot », sur larecherche.fr, La Recherche (consulté le )
- Réponse à tout !, no 227, mai 2009, p. 44.
- Bernard Heuvelmans, Sur la piste des bêtes ignorées, Paris, Plon, 1955
- « Histoire véritable du Geant Theutobocus »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?) sur viaLibri
(en) « A molecular timescale of eukaryote evolution and the rise of complex multicellular life », sur bmcevolbiol.biomedcentral.com, (consulté le ).
Voir aussi
- Société de Paléontologie (Paleontological Society), une organisation internationale dédiée à la promotion de la Paléontologie
Bibliographie
- [Buffetaut et al. 1998] Éric Buffetaut, Jean Le Loeuff, Guy Le Roux (illustr.) et Léonie Schlosser (cartographie), Les mondes disparus : atlas de la dérive des continents, Paris, Berg International, , 158 p. (ISBN 978-2-911-28914-9, OCLC 883714584).
- [Fortin, Lecointre et al. 2009] Corinne Fortin, Guillaume Lecointre (dir.) et al. (ill. Alain Bénéteau, Thomas Haessig & Dominique Visset), Guide critique de l'évolution, Paris, éd. Belin, , 571 p. (ISBN 978-2-701-14797-0, OCLC 965978019).
- [Grimaud-Hervé, Serre, Bahain et al. 2015] Dominique Grimaud-Hervé, Frédéric Serre, Jean-Jacques Bahain et al., Histoire d'ancêtres : La grande aventure de la Préhistoire, Paris IVe, Errance, coll. « Guides de la préhistoire mondiale », , 5e éd., 144 p. (ISBN 978-2-87-77-2590-3).
- [Lecointre & Guyader 2006] Guillaume Lecointre et Hervé Le Guyader, Classification phylogénétique du vivant, Paris, éd. Belin, , 559 p. (ISBN 978-2-701-14273-9, OCLC 718337975).
- [Nicholson 1877] (en) Henry Alleyne Nicholson, The ancient life-history of the earth: A comprehensive outline of the principles and leading facts of palæontological science, Edinburgh and London, William Blackwood and Sons, sur gutenberg.org (lire en ligne).
- [Steyer 2009] Sébastien Steyer (ill. Alain Bénéteau), La Terre avant les dinosaures, Paris, Belin-"Pour la Science, coll. « Bibliothéque Scientifique », , 205 p. (ISBN 978-2-701-14206-7, OCLC 698822956).
- [Tortosa 2013] Thierry Tortosa (dir.), Principes de paléontologie, Dunod, , 336 p., sur books.google.fr (ISBN 978-2-10-05-7993-8, lire en ligne).
- [Wever et al. 2010] Patrick De Wever, Bruno David, Didier Néraudeau et Jean Broutin, Paléobiosphère regards croisés des sciences de la vie et de la Terre, Paris, Muséum national d'histoire naturelle Société géologique de France Vuibert, coll. « Interactions » (no 16), , 796 p. (ISBN 978-2-711-72503-8, OCLC 732271801).
Articles connexes
- Histoire de la paléontologie
- Matériau ancien
- Paléoanthropologie
- Paléobotanique
- Paléoichnologie : le domaine de la paléontologie étudiant exclusivement les traces d'activité biologique fossilisées (ichnofossiles)
- Institut de paléontologie humaine
- Plante fossile
- Animal préhistorique
Liens externes
- Notices dans des dictionnaires ou encyclopédies généralistes :
- Ressource relative à la santé :
- Sélection de sites web sur la paléontologie dans le répertoire encyclopédique : Les Signets de la Bibliothèque nationale de France
 Portail de la paléontologie
Portail de la paléontologie  Portail de la biologie
Portail de la biologie  Portail de la géologie
Portail de la géologie