Corythosaurus
Corythosaurus (gr. "lagarto con casco") es un género representado por dos especies de dinosaurios ornitopodos hadrosáuridos, que vivieron a finales del período Cretácico, hace aproximadamente entre 77 a 75,7 millones de años, en el Campaniense, en lo que hoy es Norteamérica. Corythosaurus es clasificado como un hadrosáurido, en la subfamilias Lambeosaurinae. Estando relacionado con otros hadrosáuridos como Hypacrosaurus, Lambeosaurus y Olorotitan. Con excepción de Olorotitan todos comparten los cráneos y las crestas similares. Sin embargo, la investigación reciente ha sugerido que Olorotitan es un pariente cercano de Corythosaurus, aun sin compartir la forma de la cresta.[1]
| Corythosaurus | ||
|---|---|---|
| Rango temporal: 77 Ma - 75,7 Ma Cretácico Superior (Campaniense) | ||
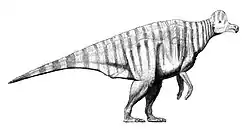 Recreación de un Corythosaurus | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Ornithischia | |
| Suborden: | Neornithischia | |
| Infraorden: | Ornithopoda | |
| (sin rango): | Iguanodontia | |
| Superfamilia: | Hadrosauroidea | |
| Familia: | Hadrosauridae | |
| Subfamilia: | Lambeosaurinae | |
| Tribu: | Corythosaurini | |
| Género: |
Corythosaurus Brown, 1914 | |
| Especie tipo | ||
|
Corythosaurus casuarius Brown,1914 | ||
| Otras Especies | ||
| ||
| Sinonimia | ||
| ||
Corythosaurus se conoce de muchos especímenes completos, incluido el holotipo casi completo encontrado por Brown en 1911. Al esqueleto holotipo solo falta la última sección de la cola y parte de las extremidades anteriores, pero se conservó con impresiones de escamas poligonales. Corythosaurus es conocido por muchos cráneos con altas crestas. Las crestas se asemejan a las crestas del casuario o a un casco corintio. La función más probable de la cresta es la vocalización. Como en un trombón, las ondas de sonido viajarían a través de muchas cámaras en la cresta, y luego se amplificarían cuando el Corythosaurus exhalara. Un espécimen de Corythosaurus se ha conservado con su última comida en su cavidad torácica. Dentro de la cavidad había restos de agujas de coníferas, semillas, ramas y frutos. Corythosaurus probablemente se alimentó de todos estos elementos. Corythosaurus tiene una longitud estimada de 9 metros, y tiene un cráneo, incluida la cresta, que mide 70,8 centímetros de alto.
Se conocen dos especies, la especie tipo y la más conocida es Corythosaurus casuarius, la otra especie es menos conocida es Corythosaurus intermedius. Ambas especies de Corythosaurus están presentes en niveles ligeramente diferentes de la Formación Dinosaur Park . Ambas coexistían con terópodos y otros ornitisquios, como Daspletosaurus, Brachylophosaurus, Parasaurolophus, Scolosaurus y Chasmosaurus.
Descripción
Benson et al. en 2012 estimaron que Corythosaurus tiene una longitud promedio de 9 metros.[2] La estimación de longitud anterior de Richard Swann Lull, publicada en 1942, encontró una longitud total ligeramente más larga de 9,4 metros, un tamaño similar al Lambeosaurus lambei , otro lambeosaurinído canadiense.[3] En 1962, Edwin H. Colbert usó modelos de dinosaurios específicos, incluido Corythosaurus, para calcular su peso. El modelo de Corythosaurus utilizado, fue modelado por Vincent Fusco parade un esqueleto montado, y supervisado por Barnum Brown. Después de la prueba, se concluyó que el peso promedio de Corythosaurus fue de 3.82 toneladas.[4] Estimaciones de tallas más recientes de Corythosaurus, publicadas en 2001, encuentran que el género está entre los hadrosáuridos más grandes, solo más pequeños que Shantungosaurus y Parasaurolophus. La longitud total del espécimen de Corythosaurus AMNH 5240 fue de 8.1 metros, con un peso de 3,0785 toneladas.[5] Proporcionalmente, el cráneo es mucho más corto y más pequeño que el de Edmontosaurus, anteriormente Trachodon, Kritosaurus o Saurolophus, pero cuando se incluye su cresta, su área superficial es casi tan grande.[6]

Corythosaurus era un herbívoro que presentaba un comportamiento gregario y se alimentaban de plantas bajas y frutos de los bosques. Su pico no tenía dientes, pero sí la mandíbula, en la que había cientos de pequeños y eran reemplazados constantemente a medida que caían. Estos eran usados para moler la vegetación de la que se alimentaba. Más de 20 cráneos se han encontrado de este dinosaurio. Como con otros lambeosáuridos, este animal tenía una cresta ósea alta, elaborada encima de su cráneo, que contenía canales nasales alargados.[7] Los canales nasales se extendieron en la cresta, primero en los bolsillos separados en los lados, luego en un solo compartimiento central y hacia adelante en el sistema respiratorio.[7] Cualquier vocalización viajaría a través de estos compartimientos elaborados, siendo amplificada.[7] Los científicos especulan que Corythosaurus podría hacer un sonido "como gritos graves como el un viento o un instrumento de latón".[7] Los sonidos serían útiles para comunicar sobre distancias largas, y podrían servir alertar a otro Corythosaurus a la presencia de alimento o a una amenaza potencial de un depredador.[7] Cuando manadas de diferentes géneros se mezclaban, su cresta, que posiblemente debe haber servido para atraer hembras y su agudo sentido olfativo debe haberle ayudado a retornar con los suyos. Alguna vez se pensó que este dinosaurio vivió sobre todo en el agua, debido al aspecto de manos y de pies palmeados.[8] Sin embargo, fue descubierto más adelante que las supuestas "membranas" era de hecho almohadillas de tejido liposo, como se encuentran en muchos mamíferos modernos.[2][9]

Un conjunto de caracteres fue indicado por Barnum Brown en 1914 para distinguir a Corythosaurus de todos los demás hadrosáuridos de Alberta, un cráneo comparativamente corto con una gran cresta en forma de casco formada por los nasales, los prefrontales y los frontales. Los nasales no están separadas al frente por los premaxilares, un pico estrecho, con una expansión frontal a una nariz alargado; y una pequeña apertura nasal.[6] En 1916 en la versión revisada, Brown expandió el conjunto de caracteres para incluir más características, un cráneo comparativamente corto con una alta cresta en forma de casco formada por nasales, prefrontales y frontales, los nasales no están separadas de la frente por los premaxilares, un pico estrecho, sección expandida en frente de las narinas alargadas, una pequeña apertura de la nariz, una fórmula vertebral de 15 cervicales, 19 dorsales, 8 sacras y más de 61 caudales, posesión de espinas dorsales de altura media, espinas caudales anteriores altas, cheurones largos, omóplatos largos, que poseen una hoja de ancho medio, un radio considerablemente más largo que el húmero, metacarpianos comparativamente cortos, un ilion anteriormente curvado, un isquion largo con una expansión terminal similar a un pie, un pubis con una lámina anterior que es corta y ampliamente expandida al final, un fémur que es más largo que la tibia; las falanges del pie son cortas, que el integumento sobre los lados y la cola se compone de escamas tuberculadas poligonales sin patrón, pero de tamaño graduado en diferentes partes del cuerpo y un vientre con hileras longitudinales de grandes escamas cónicas en forma de lapa separadas por tubérculos poligonales uniformemente grandes.[8] Los supuestos rasgos del hocico son incorrectos porque Brown confundió los premaxilares con los huesos nasales y los huesos nasales con los frontales. Actualmente se sabe que la mayoría de los rasgos postcraneales se comparten con otros lambeosaurinos.
Cráneo

Se han encontrado más de veinte cráneos de este dinosaurio. Al igual que con otros lambeosaurinos, el animal tenía una cresta ósea alta y elaborada encima de su cráneo, que contenía los pasajes nasales alargados.[7] Los pasajes nasales se extendieron por la cresta, primero en bolsas separadas en los lados, luego en una única cámara central y luego al sistema respiratorio.[2][7] El cráneo del espécimen tipo no tiene impresiones dérmicas en él. Durante la conservación, se comprimió lateralmente, por lo que ahora el ancho es aproximadamente dos tercios de lo que hubiera sido en vida. Según Brown, la compresión también provocó que las nasales se movieran donde presionaron los premaxilares. Debido a que fueron presionados en los premaxilares, los nasales habrían cerrado las narinas. Además de la compresión, el cráneo parece ser normal.[6] Al contrario de lo que Brown asumió, las áreas en cuestión eran completamente parte de los praemaxilar.
Las crestas de Corythosaurus se asemejan a las de un casuario, o un casco corintio.[2] Están formados por una combinación de los huesos premaxilar, nasales, prefrontales y frontales, como en Saurolophus, pero en lugar de proyectarse hacia atrás como una espina, se elevan para formar el punto más alto por encima de la órbita . Las dos mitades de la cresta están separadas por una sutura mediana. Por delante de la órbita, la cresta está hecha de hueso grueso.[6]

Las nasales constituyen la mayor parte de la cresta. Brown supuso que se extendían desde la punta de los picos hasta el punto más alto a lo largo de la cresta y que, a diferencia de otros géneros, las nasales se encuentran en el centro y no están separadas por un proceso premaxilar ascendente. Brown, sin embargo, confundió los premaxilares con los nasales. El hocico en realidad está formado en gran parte por ellos y separan las fosas nasales. Brown también pensó que en la parte superior y posterior de la cresta, toda la cara externa está cubierta por los frontales. Otra vez cometió un error, lo que él asumió como los frontales son, de hecho, los nasales. Las fosas nasales terminan en la parte posterior de los escamosos en un proceso corto y enganchado.[6] Los prefrontales también forman parte de la cresta, sin embargo, Brown confundió la parte baja de la rama superior del premaxilar con el prefrontal. La forma del prefrontal real, triangular, se encuentra en el lado de la base de la cresta, fue visto por Brown como parte del frontal. Los frontales reales, en gran medida internos a la estructura de la base de la cresta, no son visibles desde un lado.[10]
La boca del holotipo de Corythosaurus es estrecha. Los premaxilares forman cada uno dos largos pliegues. Los pliegues encierran conductos de aire que se extienden por los pasajes nasales hasta el frente del hocico. Allí terminan en aberturas estrechas, a veces llamadas "pseudonarinas", fosas nasales falsas. Brown los confundió con las narices reales o las fosas nasales. En realidad, se encuentran dentro de la cresta, por encima de las cuencas oculares. Como en Saurolophus, la porción expandida del premaxilar frente a la apertura del pseudonarinas es alargada, en comparación, en Kritosaurus, el pico es corto, y las pseudonarinas se extienden mucho más adelante. Al final del pico del Corythosaurus, las dos pseudonarinas se unen en una.[6] Debido a su identificación incorrecta, Brown supuso que el proceso inferior del premaxilar del holotipo era más corto que en Kritosaurus y Saurolophus, y que el proceso no se une con la lagrimal, otra diferencia de esos géneros.[6] El praemaxilar en realidad toca al lagrimal y se extiende hacia la parte posterior hasta muy atrás de la cuenca del ojo.
La mandíbula inferior del holotipo mide 66,9 centímetros de largo y 10 centímetros de profundidad. La longitud total de la cresta desde el pico hasta la punta más alta del ejemplar tipo es de 83.7 centímetros, su longitud total es de 81.2 centímetros y una altura de 70.8 centímetros.<re name=BRO14/>
Tejido Blando
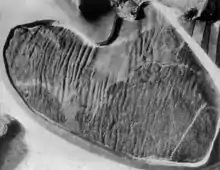
En el holotipo de C. casuarius, los lados y la cola del cuerpo están cubiertos en escamas de varios tipos. Las escamas tuberculadas poligonales, cubiertas de pequeños bultos, varían en tamaño sobre el cuerpo. Las escamas cónicas con forma de lapa solo se conservan en un pliegue de la piel preservado en la parte posterior de la tibia, pero que probablemente era parte de la región inferior del abdomen, en lugar de la pierna.[6] Separando las escamas poligonales de C. casuarius hay escamas en forma de escudo, muy juntas en filas.[11] Los tendones osificados están presentes en todas las vértebras, excepto en aquellas de la región cervical. En ninguna vértebra los tendones se extienden debajo de los procesos transversales. Cada tendón se aplana en su origen y es transversalmente ovoide en la región central, terminando en un punto redondeado.[8]
Aparte de los encontrados en Corythosaurus casuarius , se han encontrado extensas impresiones cutáneas en Edmontosaurus annectens y se ha encontrado una tegumentaria notable en Brachylophosaurus canadensis, Gryposaurus notabilis , Parasaurolophus walkeri , Lambeosaurus magnicristatus , L. lambei , Saurolophus angustirsotris y en ornitópodos no identificados. De estos, L. lambei, C. casuarius, G. notabilis, P. walkeri y S. angustirsotris han conservado escamas poligonales. Las escamas en L. lambei, S. angustirostris y C. casuarius son todos similares. Corythosaurus es uno de los pocos hadrosáuridos que han conservado las impresiones de la piel en las extremidades posteriores y los pies. Un estudio en 2013 mostró que entre los hadrosáuridos, Saurolophus angustirostris conservó la mejor y más completa integumentación de pies y extremidades, aunque otras especies como S. osborni, Edmontosaurus annectens y Lambeosaurus lambei (= L. clavinitialis) comparten una buena cantidad de tejido conservado en las regiones.[11]
Descubrimiento e investigación

El primer espécimen, AMNH 5240, fue descubierto en 1911 por Barnum Brown en Río Red Deer, Alberta , Canadá y asegurado por él en el otoño de 1912.[12][6] Además del esqueleto casi completo, el hallazgo fue notable porque las impresiones de gran parte de la piel de la criatura también sobrevivieron.[6] El espécimen provino del Grupo Belly River.[6] La parte izquierda o la parte inferior del esqueleto se conserva en arcilla carbonosa, por lo que es difícil exponer la piel.[6] El esqueleto se encontró articulado, y solo falta en los últimos 0,61 metros de la cola y las extremidades anteriores.[6] Ambas escápulas y coracoides se conservan en posición, pero el resto de las extremidades anteriores se han perdido, a excepción de falanges y trozos de húmero, cúbitos y radios. Aparentemente, las extremidades anteriores restantes fueron desgastadas o erosionadas.[6] Las impresiones de la epidermis se conservaron cubriendo una gran parte de los esqueletos que delinean, y muestra la forma del cuerpo.[6] Otro espécimen, AMNH 5338, fue encontrado en 1914 por Brown y Peter Kaisen. Ambos especímenes ahora se encuentran en el Museo Americano de Historia Natural en sus poses de muerte originales.[13]
La especie tipo, Corythosaurus casuarius, fue nombrada por Barnum Brown en 1914, basándose en el primer espécimen recogido por él en 1912. AMNH 5240 es, por lo tanto, el holotipo . En 1916, el autor original, Brown, publicó una descripción más detallada que también se basó en AMNH 5338, cuyo espécimen es por lo tanto el plesiotipo . El Corythosaurus es uno de los muchos lambeosaurinídos que poseen crestas, y fue la cresta la que le dio su nombre al Corythosaurus . El nombre genérico Corythosaurus se deriva del griego κόρυθος, korythos, "casco corintio", y significa "lagarto con casco".[13] El nombre específico casuarius se refiere al casuario, un ave con una cresta craneal similar. El binomio completo de Corythosaurus casuarius significa "reptil tipo casuario, con una cresta de casco corintio".[14]

Los dos ejemplares mejor conservados de Corythosaurus encontrados, por Charles H. Sternberg en 1912, se perdieron el 6 de diciembre de 1916 cuando eran llevados por el SS Mount Temple al Reino Unido , durante la Primera Guerra Mundial . Los enviaban a Arthur Smith Woodward, un paleontólogo del Museo Británico de Historia Natural en Inglaterra , cuando el buque mercante que los transportaba fue hundido por el mercante asaltante alemán SMS Möwe en medio del océano.[15]
_(20135543774).jpg.webp)
Había antiguamente hasta siete especies descritas, incluyendo C. casuarius, C. bicristatus Parks 1935, C. brevicristatus Parks 1935, C. excavatus Gilmore 1923, C. frontalis Parks 1935, y C. intermedius Parks 1923. En 1975 Peter Dodson estudió las diferencias entre los cráneos y las crestas de diferentes especies de dinosaurios lambeosaurinos. Descubrió que las diferencias de tamaño y forma pueden estar relacionadas con el sexo y la edad del animal. Ahora solo se reconoce con certeza una especie, C. casuarius,[16] aunque C. intermedius ha sido reconocido como válido en algunos estudios. Se basa en el espécimen ROM 776, un cráneo encontrado por Levi Sternberg en 1920 y fue nombrado por William Parks en 1923, que originalmente lo había llamado Stephanosaurus intermedius a principios de ese año.[17] El nombre específico de C. intermedius se deriva de su posición intermedia aparente de acuerdo con Parks.[18][19][20] C. intermedius vivió un poco más tarde en el Campaniano que C. casuarius y las dos especies no son idénticas, lo que apoyó la separación de ellas en un estudio de 2009.[21]
Clasificación
Originalmente, Brown refirió a Corythosaurus como un miembro de la familia Trachodontidae[6],ahora Hadrosauridae.[2] Dentro de Trachodontidae estaban las subfamilias Trachodontinae y Saurolophinae. Brown clasificó Hadrosaurus, Trachodon, Claosaurus y Kritosaurus en Trachodontinae y Corythosaurus , Stephanosaurus y Saurolophus en Saurolophinae.[6]
Más tarde, Brown revisó la filogenia de Corythosaurus , y descubrió que estaba estrechamente relacionada, y posiblemente era el ancestro de Hypacrosaurus. Las únicas diferencias que encontró entre ellos fueron el desarrollo de las vértebras y las proporciones de las extremidades.[8] Durante un estudio de huesos ilíacos de dinosaurios en la década de 1920, Alfred Sherwood Romer propuso que las dos órdenes de dinosaurios podrían haber evolucionado por separado, y que las aves, según la forma y las proporciones de su ilion, podrían ser ornitisquios especializados. Usó a Tyrannosaurus y Corythosaurus como modelos base para analizar qué teoría era más probable de ser cierta. Descubrió que, aunque se piensa que las aves son saurisquios, es muy plausible que hayan evolucionado su musculatura y anatomía pélvica específica si evolucionaron de ornitisquios como Corythosaurus.[22] Sin embargo, a pesar de que Corythosaurus se asemeja más a aves modernas en estructura pélvica que a los saurisquios, ahora se cree que las aves han descendido de estos últimos.[2]
Corythosaurus se clasifica actualmente como un hadrosáurido , en la subfamilia Lambeosaurinae. Se relaciona con otros hadrosaurios como Hypacrosaurus , Lambeosaurus y Olorotitan que con la excepción de Olorotitan, todos comparten cráneos y crestas similares. Sin embargo, una investigación publicada en 2003 ha sugerido que, aunque posee una cresta única, Olorotitan es el pariente más cercano conocido de Corythosaurus.[1] Benson et al. en 2012 encontraron que Corythosaurus estaba estrechamente relacionado con Velafrons , Nipponosaurus e Hypacrosaurus y que formaban un grupo de lambeosaurines con cresta de abanico.[2]
En 2014, se publicó un estudio que incluye la descripción de Zhanghenglong en la revista PLOS ONE . El estudio incluyó un cladograma casi completo de las relaciones de los hadrosauroideos , que incluyen Corythosaurus como el lambeosaurínido más derivado, como taxón hermano de Hypacrosaurus.[23]
Filogenia
El siguiente cladograma es una versión simplificada del estudio de 2014 que incluye solo a los Lambeosaurini.[23]

| Lambeosaurini |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiología
Las comparaciones entre los anillos esclerales de Corythosaurus con las aves y reptiles modernos sugieren que puede haber sido catemeral, es decir activo durante todo el día en intervalos cortos.[9] El sentido del oído en hadrosáuridos, específicamente estudiado en Lophorhothon, también parece haber estado muy desarrollado debido a una extremidad del conducto coclear cerrada alargado.[24] La presencia de un estribo delgado, un hueso de la oreja que es similar a una vara en los reptiles, combinado con un tímpano grande implica la existencia de un oído medio sensible.[9] posible que los oídos de los hadrosáuridos sean lo suficientemente sensibles para detectar tanto sonido como un cocodrilo moderno.[24]
Función de la Cresta
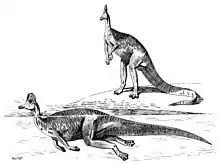
Las estructuras internas de la cresta de Corythosaurus son bastante complejas, lo que hace posible la utilización para realizar una llamada que podría utilizarse para advertir o atraer pareja. Los conductos nasales de Corythosaurus, así como Hypacrosaurus y Lambeosaurus tienen forma de S, y Parasaurolophus solo posee tubos en forma de U.[24] Cualquier vocalización viajaría a través de estas elaboradas cámaras, y probablemente se amplificaría.[2][7] Los científicos especulan que el Corythosaurus podría emitir gritos agudos y bajos "como un instrumento de viento o de latón",[7] como un trombón.[13] Los sonidos podrían servir para alertar a otros Corythosaurus a la presencia de comida o una amenaza potencial de un depredador.[7] Los pasajes nasales emiten sonidos de baja frecuencia cuando Corythosaurus exhala. Las crestas individuales producirían diferentes sonidos, por lo que es probable que cada especie de lambeosaurine haya tenido un sonido único.[13] Sin embargo, a pesar de que el rango para diferentes pasajes nasales de lambeosaurine varía, todos ellos probablemente emitieron sonidos de tono bajo. Esto podría deberse a que los sonidos bajos, por debajo de 400 hertcios, recorren una distancia establecida en cualquier entorno, mientras que los sonidos más alto, más de 400 Hz, tienen una mayor dispersión en la distancia recorrida.[24]
Cuando se describieron por primera vez, se creía que los hadrosáuridos con cresta eran acuáticos,[13] una evaluación basada incorrectamente en las correas que ahora se conoce como relleno.[2][8] La teoría era que los animales podían nadar en las profundidades del agua y usar la cresta para almacenar aire para respirar. Sin embargo, ahora se ha demostrado que la cresta no tenía agujeros en el extremo, y la presión del agua a incluso 3 metros sería demasiado grande para que los pulmones puedan inflarse.[13]
Ontogenia

Corythosaurus casuarius es uno de los pocos lambeosaurinídes, junto con Lambeosaurus lambei, Hypacrosaurus stebingeri y H. altispinus que presentan juveniles fosilizados asignados. Los juveniles son más difíciles de asignar a las especies, porque a una edad temprana carecen de las crestas distintivas más grandes de los adultos. A medida que envejecen, las crestas de los lambeosaurinos tienden a crecer y se vuelven más prominentes en la madurez. En la Formación Dinosaur Park, se han encontrado más de cincuenta especímenes articulados, procedentes de muchos géneros diferentes. Entre ellos, los juveniles son difíciles de identificar a nivel de especie. Anteriormente, se reconocieron cuatro géneros y trece especies del área de la formación, cuando los paleontólogos utilizaron las diferencias en tamaño y forma de cresta para diferenciar los taxones. Los especímenes más pequeños fueron identificados como Tetragonosaurus , ahora visto como un sinónimo de Procheneosaurus, y los esqueletos más grandes se llamaron Corythosaurus o Lambeosaurus, un adulto fue identificado como Parasaurolophus. Los pequeños lambeosaurinos de la Formación Horseshoe Canyon fueron derivados a Cheneosaurus.[25]

Corythosaurus comenzó a desarrollar su cresta cuando tenía la mitad del tamaño de adulto, pero los juveniles de Parasaurolophus comenzaron a desarrollar las crestas cuando solo tenían el 25% de los adultos. El Corythosaurus juvenil , junto con los adultos, tenía una fontanela premaxilar nasal. Corythosaurus jóvenes y adultos son similares a Lambeosaurus e Hypacrosaurus pero diferentes a Parasaurolophus en que las suturas del cráneo son sinuosas, y no lisas y rectas. Esta característica ayuda a diferenciara los parasaurolofínidos de lambeosaurinidos. Generalmente, las crestas de juveniles de lambeosaurinídos como Corythosaurus, Lambeosaurus e Hypacrosaurus stebingeri, parasaurolofínidos como Parasaurolophus y primitivos lambeosauridos como Kazaklambia son bastante parecidos, aunque se pueden usar otras características para distinguirlos.[26]
El trabajo de Dodson de 1975 reconoció que había muchos menos taxones presentes en Alberta.[16][25] Tetragonosaurus resultó ser juveniles de Corythosaurus o Lambeosaurus . T. erectofrons fue asignado a Corythosaurus basado en gran medida en información biométrica. El único espécimen no típico de Tetragonosaurus , asignado a T. erectofrons, se encontró más tarde referible a Hypacrosaurus , aunque se descubrió que el holotipo de la especie era asignable a Corythosaurus.[25]
Dieta
Corythosaurus era un ornitópodo, y por lo tanto un herbívoro. Benson et al. en 2012 se dieron cuenta de que el pico de Corythosaurus era poco profundo y delicado, y concluyeron que debe haber sido utilizado para alimentarse de vegetación blanda. Basado en el clima del Cretácico superior, supusieron que el Corythosaurus habría sido un ramoreador selectivo, comiendo solo las frutas más jugosas y las hojas más jóvenes.[2] Las muestras de Corythosaurus que se han conservado con su última comida en su cavidad torácica. Dentro de la cavidad había restos de agujas de coníferas , semillas, ramitas y frutos, lo que significa que el Corythosaurus probablemente se alimentó de todos estos.[13]
En la cultura popular
Corythosaurus ha aparecido llegó al universo cinematográfico por la película de Parque jurásico 3 en la que sólo aparece un grupo de estos animales junto a Parasaurolophus apareciendo en unos pocas escenas durante la persecución por parte de unos letales Velociraptores. También hace aparición en la serie infantil de Dino dan de 2009 con un color de piel similar al de un pez payaso.
Paleoecología
Se han encontrado fósiles en la formación Oldman superior y en la formación Dinosaur Park de Canadá . La formación Oldman data del Campaniano , hace unos 77,5 a 76,5 millones de años,[21] y la formación Dinosaur Park data de hace 76,6 a 74,8 millones de años.[21][27] Corythosaurus vivió entre 77 a 75.7 millones de años atrás. En la formación Dinosaur Park, C. casuarius vivió entre 76,6 y 75,9 millones de años atrás, con C. intermedius viviendo de 75,8 a 75,7 millones de años atrás. En la formación Oldman C. casuarius , la única especie de Corythosaurus de los depósitos, vivió alrededor de 77 a 76.5 millones de años atrás.[21] El espécimen de holotipo era claramente un cadáver que había flotado en una playa, ya que las conchas del género Unio, los huesos desgastados por el agua y una tortuga baenida se conservaban alrededor.[8] Corythosaurus probablemente vivió en un bosque de bosque, y de vez en cuando pudo vagar por áreas pantanosas.[2]

Se conoce una fauna limitada de la sección superior de la Formación Oldman, y Corythosaurus casuarius y C. intermedius se encuentran entre los taxones. También de la sección de la formación se encuentran los terópodos Daspletosaurus y Saurornitholestes, los hadrosáuridos Brachylophosaurus, Gryposaurus y Parasaurolophus , el anquilosáuridos Scolosaurus y los ceratópsidos Coronosaurus y Chasmosaurus . Se conocen otros géneros en la formación, pero no persisten en la sección superior de la formación, y por lo tanto no son contemporáneos de Corythosaurus.[21]
Corythosaurus casuarius está muy extendido en toda la unidad inferior de la Formación Dinosaur Park.[28] En ella, se descubrió que Corythosaurus estaba estrechamente relacionado con el ceratósido Centrosaurus apertus . Su asociación se encontró en las formaciones Dinosaur Park, Judith River y Mesaverde , y también en el bajio Wind River y Wheatland County .[29] Corythosaurus vivió junto a muchos otros herbívoros gigantes, como los hadrosáuridos Gryposaurus y Parasaurolophus , los ceratopsidos Centrosaurus y Chasmosaurus, y anquilosaurianos Scolosaurus, Edmontonia' y Dyoplosaurus en las etapas más tempranas de la formación, Dyoplosaurus, Panoplosaurus[21] y Euoplocephalus en la edad media, y Euoplocephalus solo en etapas posteriores de la formación. Los estudios de la anatomía de la mandíbula y la mecánica de estos dinosaurios sugieren que probablemente todos ocuparon nichos ecológicos ligeramente diferentes con el fin de evitar la competencia directa por la comida en un espacio ecológico tan concurrido.[28] Los únicos grandes depredadores conocidos de los mismos niveles de la formación como Corythosaurus son los tiranosáuridos Gorgosaurus libratus y una especie anónima de Daspletosaurus.[21]
Thomas M. Lehman ha observado que el Corythosaurus no se ha descubierto fuera del sur de Alberta a pesar de que es uno de los dinosaurios Juditiano más abundantes en la región. Grandes herbívoros como los hadrosaurios que vivían en América del Norte durante el Cretácico superior tenían "rangos geográficos notablemente pequeños" a pesar de su gran tamaño corporal y alta movilidad. Esta distribución restringida contrasta fuertemente con las faunas modernas de mamíferos cuyos grandes rangos de herbívoros "típicamente ... abarcan gran parte de un continente".[29]
Referencias
- Godefroit, Pascal; Bolotsky, Yuri; Alifanov, Vladimir (2003). «A remarkable hollow-crested hadrosaur from Russia: an Asian origin for lambeosaurines». Comptes Rendus Palevol 2: 143-151. doi:10.1016/S1631-0683(03)00017-4.
- Benson, R.B.J.; Brussatte, M.; Xu, X. (2012). Prehistoric Life. London: Dorling Kindersley. pp. 344–345. ISBN 978-0-7566-9910-9.
- Lull, R.S.; Wright, N.E. (1942). «Hadrosaurian Dinosaurs of North America». Geological Society of America Special Papers 40: 1-272. doi:10.1130/SPE40-p1.
- Colbert, E.H. (1962). «The Weight of Dinosaurs». American Museum Novitates 2076: 1-16.
- Seebacher, F. (2001). «A New Method to Calculate Allometric Length-Mass Relationships of Dinosaurs». Journal of Vertebrate Paleontology 21 (1): 51-60. doi:10.1671/0272-4634(2001)021[0051:ANMTCA]2.0.CO;2.
- Brown, B. (1914). «Corythosaurus casuarius, a New Crested Dinosaur from the Belly River Cretaceous, with Provisional Classification of the Family Trachodontidae». American Museum of Natural History Bulletin 33: 559-565.
- "Corythosaurus." In: Dodson, Peter & Britt, Brooks & Carpenter, Kenneth & Forster, Catherine A. & Gillette, David D. & Norell, Mark A. & Olshevsky, George & Parrish, J. Michael & Weishampel, David B. The Age of Dinosaurs. Publications International, LTD. p. 137. ISBN 0-7853-0443-6.
- Brown, B. (1916). «Corythosaurus casuarius: Skeleton, Musculature and Epidermis». American Museum of Natural History Bulletin 38: 709-715.
- Benson, R.B.J.; Brussatte, M.; Xu, X. (2012). Prehistoric Life. London: Dorling Kindersley. pp. 344–345. ISBN 978-0-7566-9910-9.
- Schmitz, L. and Motani, R. (2011). «Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology». Science 332 (6030): 705-8. PMID 21493820. doi:10.1126/science.1200043.
- Weishampel, D.B., Dodson, P., Osmólska, H., & Hilton, Richard P., 2004, The Dinosauria. University of California Press. p. 450
- Bell, P. R. (2012). «Standardized terminology and potential taxonomic utility for hadrosaurid skin impressions: A case study for Saurolophus from Canada and Mongolia». PLoS ONE 7 (2): e31295. PMC 3272031. PMID 22319623. doi:10.1371/journal.pone.0031295.
- Lowell Dingus and Mark Norell, 2011 Barnum Brown: The Man Who Discovered Tyrannosaurus rex, University of California Press, p. 143
- Norrell, M.; Gaffney, E.S.; Dingus, L. (2000). Discovering Dinosaurs: Evolution, Extinction, and the Lessons of Prehistory. London: University of California Press. pp. 35, 158–159. ISBN 0-520-22501-5.
- «DinoGuide: Corythosaurus casuarius». Dinosaurs in Their Time. Carnegie Museum of Natural History. Archivado desde el original el 10 de agosto de 2014. Consultado el 27 de julio de 2014.
- Tanke, D.H. & Carpenter, K. (2001). Mesozoic Vertebrate Life. Indiana University Press. pp. 280–328. ISBN 0-253-33907-3.
- Dodson, P. (1975). «Taxonomic implications of relative growth in lambeosaurine dinosaurs». Systematic Zoology 24 (1): 37-54. JSTOR 2412696. doi:10.2307/2412696.
- Parks, W.A. (1923). «New species of crested trachodont dinosaur». Bulletin of the Geological Society of America 34: 130.
- Parks, W.A. (1923). «Corythosaurus intermedius, a new species of trachodont dinosaur». University of Toronto Studies, Geological Series 15: 5-57.
- Prieto-Márquez, A.; Vecchia, F.M.D.; Gaete, R.; Galobart, À. (2013). «Diversity, Relationships, and Biogeography of the Lambeosaurine Dinosaurs from the European Archipelago, with Description of the New Aralosaurin Canardia garonnensis». En Dodson, Peter, ed. PLoS ONE 8 (7): e69835. PMC 3724916. PMID 23922815. doi:10.1371/journal.pone.0069835.
- Campione, N. E.; Brink, K. S.; Freedman, E. A.; McGarrity, C. T.; Evans, D. C. (2013). «Glishades ericksoni, an indeterminate juvenile hadrosaurid from the Two Medicine Formation of Montana: implications for hadrosauroid diversity in the latest Cretaceous (Campanian-Maastrichtian) of western North America». Palaeobiodiversity and Palaeoenvironments 93: 65-75.
- Arbour, V. M.; Burns, M. E.; Sissons, R. L. (2009). «A redescription of the ankylosaurid dinosaur Dyoplosaurus acutosquameus Parks, 1924 (Ornithischia: Ankylosauria) and a revision of the genus». Journal of Vertebrate Paleontology 29 (4): 1117-1135. doi:10.1671/039.029.0405.
- Romer, A.S. (1923). «The Ilium in Dinosaurs and Birds». Bulletin of the AMNH 48 (5): 141-145.
- Xing, H.; Wang, D.; Han, F.; Sullivan, C.; Ma, Q.; He, Y.; Hone, D. W. E.; Yan, R.; Du, F.; Xu, X. (2014). «A New Basal Hadrosauroid Dinosaur (Dinosauria: Ornithopoda) with Transitional Features from the Late Cretaceous of Henan Province, China». PLoS ONE 9 (6): e98821. PMC 4047018. PMID 24901454. doi:10.1371/journal.pone.0098821.
- Weishampel, D.B. (1981). «Acoustic Analysis of Vocalization of Lambeosaurine Dinosaurs (Reptilia: Ornithischia)». Paleobiology 7 (2): 252-261. JSTOR 2400478. Archivado desde el original el 6 de octubre de 2014.
- Evans, D.C.; Forster, C.A.; Reiz, R.R. (2005). «The Type Specimen of Tetragonosaurus erectofrons (Ornithischia: Hadrosauridae) and the Identification of Juvenile Lambeosaurines». En Currie, P.J. & Koppelhus, Eva, B., ed. Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press. pp. 349–363. ISBN 0-253-34595-2.
- Farke, A. A.; Chok, D. J.; Herrero, A.; Scolieri, B.; Werning, S. (2013). «Ontogeny in the tube-crested dinosaur Parasaurolophus (Hadrosauridae) and heterochrony in hadrosaurids». PeerJ 1: e182. PMC 3807589. PMID 24167777. doi:10.7717/peerj.182.
- Currie, P.J. & Koppelhus, E.B. (2005). Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press. pp. 54–435. ISBN 0-253-34595-2.
- Mallon, Jordan C.; Evans, David C.; Ryan, Michael J.; Anderson, Jason S. (2012). «Megaherbivorous dinosaur turnover in the Dinosaur Park Formation (upper Campanian) of Alberta, Canada». Palaeogeography, Palaeoclimatology, Palaeoecology. 350–352: 124-138. doi:10.1016/j.palaeo.2012.06.024.
- Tanke, D.H. & Carpenter, K. (2001). Mesozoic Vertebrate Life. Indiana University Press. pp. 280–328. ISBN 0-253-33907-3.
Véase también
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Wikimedia Commons alberga una categoría multimedia sobre Corythosaurus.
Wikimedia Commons alberga una categoría multimedia sobre Corythosaurus. Wikispecies tiene un artículo sobre Corythosaurus.
Wikispecies tiene un artículo sobre Corythosaurus.- Corythosaurus dentro de Lambeosaurinae en The Thescelsosaurus (en inglés)
- Significado y pronunciación del nombre (en inglés)
- CIENCIA AL CUBO: Podcast sobre la cresta de los dinosaurios
| Control de autoridades |
|
|---|
 Datos: Q131078
Datos: Q131078- Multimedia: Corythosaurus / Q131078
- Especies: Corythosaurus