Solénoïde (domaine protéique)
Un domaine protéique en solénoïde est un type de structure hautement conservé fréquemment rencontré dans les protéines. Il s'agit d'une chaîne de motifs structurels répétés à l'identique ou pratiquement à l'identique. On parle souvent simplement de « répétitions ». De telles structures sont extrêmement courantes parmi les protéines, bien que leur nombre exact ne soit pas connu[1].
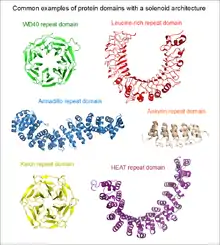
Répétitions de motifs dans les protéines
Dans une protéine, une répétition est un bloc de séquence peptidique reproduit plus d'une fois dans l'ensemble de la séquence de la protéine, à l'identique ou de manière très semblable. D'une manière générale, les motifs répétés de moins de 10 résidus d'acides aminés ne sont pas significatifs et peuvent être tout à fait désordonnés, tandis que des motifs de 30 à 40 résidus ont toutes les chances de former une structure protéique particulière, et sont souvent des indicateurs d'une structure en solénoïde dans la protéine.
On trouve des répétitions désordonnées par exemple parmi les motifs de sept résidus de la sous-unité RPB1 de l'ARN polymérase II[2], dans la β-caténine ou encore dans le motif linéaire de liaison de l'axine dans la protéine APC[3]. On trouve des répétitions ordonnées de séquences courtes par exemple dans les répétitions de trois résidus du collagène ou encore dans les répétitions pentapeptidiques formant les structures en hélice β.
Solénoïdes ouverts et solénoïdes fermés
Compte tenu de leur mode de construction, les domaines protéiques en solénoïdes ne peuvent prendre qu'un nombre limité de formes. Il en existe deux types principaux : les solénoïdes linéaires, ou ouverts, généralement plus ou moins hélicoïdaux, et les solénoïdes circulaires, ou fermés.
Solénoïdes linéaires (ouverts)
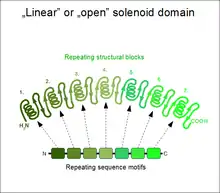
Lorsque les deux répétitions terminales du solénoïde n'interagissent pas entre elles, on a affaire à une structure dite ouverte, ou linéaire. De tels solénoïdes présentent généralement une forme en bâtonnet ou en croissant. Le nombre de répétitions qui les constituent va de 2 à plus de 50. Un avantage évident de ce type de structure est que les extrémités N-terminale et C-terminale peuvent être allongées par de nouvelles répétitions ou raccourcies par élimination de certaines d'entre elles sans que cela ait un impact significatif sur la stabilité de l'ensemble du domaine[4]. Ce type de domaine est extrêmement courant parmi les segments extracellulaires de récepteurs ou de protéines d'adhérence cellulaire. Ce sont par exemple la répétition EGF, la répétition cadhérine, la répétition riche en leucine, la répétition HEAT, la répétition ankyrine, la répétition armadillo, répétition tétratricopeptide, etc. Lorsqu'un domaine solénoïde linéaire participe à une interaction protéine-protéine, le site de liaison au ligand est fréquemment constitué d'au moins trois répétitions. Les unités répétées ne peuvent ainsi généralement pas assurer seules les fonctions de l'ensemble du domaine.
Solénoïdes circulaires (fermés)
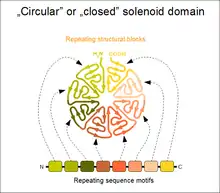
Lorsque les deux répétitions terminale du solénoïde interagissent entre elles, il se forme une structure topologiquement fermée et compacte. De tels domaines présentent généralement une symétrie circulaire, contrairement aux solénoïdes ouverts qui présentent seulement une symétrie de translation, et peuvent prendre une forme de roue. Compte tenu des limitations de cette structure, le nombre d'éléments répétés n'est pas arbitraire. Dans le cas des répétitions WD40, le nombre de répétitions peut aller de 4 à 10, le plus souvent entre 5 et 7[5]. Les motifs Kelch, les tonneaux β et le trèfle β sont d'autres exemples de ce type de structures. Les solénoïdes fermés fonctionnent souvent comme des modules d'interaction protéine-protéine : toutes les unités répétées doivent être généralement présentes afin de former le site de liaison au ligand si celui-ci est situé au centre ou dans l'axe du solénoïde.
Supra-domaines de motifs répétés
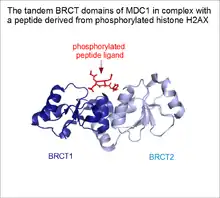
Comme souvent en biologie, il existe différents cas limites entre structures en solénoïde et les autres types de domaines protéiques. Les protéines qui contiennent des répétitions en tandem de domaines ordinaires sont très courantes chez les eucaryotes. Bien que certains d'entre eux sont parfaitement capables de se replier de manière indépendante, certains d'entre eux peuvent se lier ensemble et prendre une disposition fixe dans la protéine. De tels modules supra-domaines sont susceptibles d'assurer des fonctions dont leurs constituants individuels sont incapables[6]. Un exemple bien connu est le tandem de domaines BRCT de la protéine BRCA1[7]. Bien que certains domaines BRCT individuels soient présents dans certaines protéines comme les ADN ligases, ces domaines BRCT en tandem ont développé une fonction nouvelle dans la mesure où ils assurent la fonction de motif de liaison à des ligands peptidiques phosphorylés[8],[9]. Dans le cas des protéines BRCA1 et MDC1 (en), la fente de liaison au peptide se trouve dans la crevasse formée par la jonction des deux domaines. C'est la raison pour laquelle les constituants individuels de ce supra-domaine ne peuvent assurer seuls la liaison au ligand. Pour cette raison, les domaines BRCT peuvent être vus comme une forme de solénoïde linéaire simple.
Notes et références
- (en) Miguel A. Andrade, Carolina Perez-Iratxeta et Chris P. Ponting, « Protein Repeats: Structures, Functions, and Evolution », Journal of Structural Biology, vol. 134, nos 2-3, , p. 117-131 (PMID 11551174, DOI 10.1006/jsbi.2001.4392, lire en ligne)
- (en) Peter A. Meyer, Ping Ye, Mincheng Zhang, Man-Hee Suh et Jianhua Fu, « Phasing RNA Polymerase II Using Intrinsically Bound Zn Atoms: An Updated Structural Model », Structure, vol. 14, no 6, , p. 973-982 (PMID 16765890, DOI 10.1016/j.str.2006.04.003, lire en ligne)
- (en) Jing Liu, Yi Xing, Thomas R. Hinds, Jie Zheng et Wenqing Xu, « The Third 20 Amino Acid Repeat Is the Tightest Binding Site of APC for β-Catenin », Journal of Molecular Biology, vol. 360, no 1, , p. 133-144 (PMID 16753179, DOI 10.1016/j.jmb.2006.04.064, lire en ligne)
- (en) Lisa N. Kinch et Nick V. Grishin, « Evolution of protein structures and functions », Current Opinion in Structural Biology, vol. 12, no 3, , p. 400-408 (PMID 12127461, DOI 10.1016/S0959-440X(02)00338-X, lire en ligne)
- (en) Cammy K.-M. Chen, Nei-Li Chan et Andrew H.-J. Wang, « The many blades of the β-propeller proteins: conserved but versatile », Trends in Biochemical Sciences, vol. 36, no 10, , p. 553-561 (PMID 21924917, DOI 10.1016/j.tibs.2011.07.004, lire en ligne)
- (en) Christine Vogel, Carlo Berzuini, Matthew Bashton, Julian Gough et Sarah A. Teichmann, « Supra-domains: Evolutionary Units Larger than Single Protein Domains », Journal of Molecular Biology, vol. 336, no 3, , p. 809-823 (PMID 15095989, DOI 10.1016/j.jmb.2003.12.026, lire en ligne)
- (en) Xiaochun Yu, Claudia Christiano Silva Chini, Miao He, Georges Mer et Junjie Chen, « The BRCT Domain Is a Phospho-Protein Binding Domain », Science, vol. 302, no 5645, , p. 639-642 (PMID 14576433, DOI 10.1126/science.1088753, lire en ligne)
- (en) Zi-Zhang Sheng, Yu-Qi Zhao et Jing-Fei Huang, « Functional Evolution of BRCT Domains from Binding DNA to Protein », Evolutionary Bioinformatics, vol. 7, , p. 87-97 (PMID 21814458, PMCID 3140412, DOI 10.4137/EBO.S7084, lire en ligne)
- (en) Charles Chung Yun Leung et J. N. Mark Glover, « BRCT domains: Easy as one, two, three », Cell Cycle, vol. 10, no 15, , p. 2461-2470 (PMID 21734457, PMCID 3180187, DOI 10.4161/cc.10.15.16312, lire en ligne)
 Portail de la biochimie
Portail de la biochimie  Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire