RUNX2
Runt-related transcription factor 2 (RUNX2) also known as core-binding factor subunit alpha-1 (CBF-alpha-1) is a protein that in humans is encoded by the RUNX2 gene. RUNX2 is a key transcription factor associated with osteoblast differentiation.




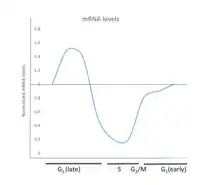
It has also been suggested that Runx2 plays a cell proliferation regulatory role in cell cycle entry and exit in osteoblasts, as well as endothelial cells. Runx2 suppresses pre-osteoblast proliferation by affecting cell cycle progression in the G1 phase.[6] In osteoblasts, the levels of Runx2 is highest in G1 phase and is lowest in S, G2, and M.[5] The comprehensive cell cycle regulatory mechanisms that Runx2 may play are still unknown, although it is generally accepted that the varying activity and levels of Runx2 throughout the cell cycle contribute to cell cycle entry and exit, as well as cell cycle progression. These functions are especially important when discussing bone cancer, particularly osteosarcoma development, that can be attributed to aberrant cell proliferation control.
Function
Osteoblast differentiation
This protein is a member of the RUNX family of transcription factors and has a Runt DNA-binding domain. It is essential for osteoblastic differentiation and skeletal morphogenesis. It acts as a scaffold for nucleic acids and regulatory factors involved in skeletal gene expression. The protein can bind DNA both as a monomer or, with more affinity, as a subunit of a heterodimeric complex. Transcript variants of the gene that encode different protein isoforms result from the use of alternate promoters as well as alternate splicing.
The cellular dynamics of Runx2 protein are also important for proper osteoblast differentiation. Runx2 protein is detected in preosteoblasts and the expression is upregulated in immature osteoblasts and downregulated in mature osteoblasts. It is the first transcription factor required for determination of osteoblast commitment, followed by Sp7 and Wnt-signaling. Runx2 is responsible for inducing the differentiation of multipotent mesenchymal cells into immature osteoblasts, as well as activating expression of several key downstream proteins that maintain osteoblast differentiation and bone matrix genes.
Knock-out of the DNA-binding activity results in inhibition of osteoblastic differentiation. Because of this, Runx2 is often referred to as the master regulator of bone.[7]
Cell cycle regulation
In addition to being the master regulator of osteoblast differentiation, Runx2 has also been shown to play several roles in cell cycle regulation. This is due, in part, to the fact that Runx2 interacts with many cellular proliferation genes on a transcription level, such as c-Myb and C/EBP,[5] as well as p53/[7] These functions are critical for osteoblast proliferation and maintenance. This is often controlled via oscillating levels of Runx2 within throughout cell cycle due to regulated degradation and transcriptional activity.
Oscillating levels of Runx2 within the cell contribute to cell cycle dynamics. In the MC3T3-E1 osteoblast cell line, Runx2 levels are a maximum during G1 and a minimum during G2, S, and mitosis.[5] In addition, the oscillations in Runx2 contribute to G1-related anti-proliferative function.[8] It has also been proposed that decreasing levels of Runx2 leads to cell cycle exit for proliferating and differentiating osteoblasts, and that Runx2 plays a role in mediating the final stages of osteoblast via this mechanism.[9] Current research posits that the levels of Runx2 serve various functions.
In addition, Runx2 has been shown to interact with several kinases that contribute to facilitate cell-cycle dependent dynamics via direct protein phosphorylation. Furthermore, Runx2 controls the gene expression of cyclin D2, D3, and the CDK inhibitor p21(cip1) in hematopoietic cells. It has been shown that on a molecular level, Runx associates with the cdc2 partner cyclin B1 during mitosis.[10] The phosphorylation state of Runx2 also mediates its DNA-binding activity. The Runx2 DNA-binding activity is correlated with cellular proliferation, which suggests Runx2 phosphorylation may also be related to Runx2-mediated cellular proliferation and cell cycle control. To support this, it has been noted that Runx is phosphorylated at Ser451 by cdc2 kinase, which facilitates cell cycle progression through the regulation of G2 and M phases.[10]

Pathology
Cleidocranial dysplasia
Mutations in Runx2 are associated with the disease Cleidocranial dysostosis. One study proposes that this phenotype arises partly due to the Runx2 dosage insufficiencies. Because Runx2 promotes exit from the cell cycle, insufficient amounts of Runx2 are related to increased proliferation of osteoblasts observed in patients with cleidocranial disostosis.[11]
Osteosarcoma
Variants of Runx2 have been associated with the osteosarcoma phenotype.[5] Current research suggests that this is partly due to the role of Runx2 in mitigating the cell cycle.[6] Runx2 plays a role as a tumor suppressor of osteoblasts by halting cell cycle progression at G1.[5] Compared to normal osteoblast cell line MC3T3-E1, the oscillations of Runx2 in osteosarcoma ROS and SaOS cell lines are aberrant when compared to the oscillations of Runx2 levels in normal osteoblasts, suggesting that deregulation of Runx2 levels may contribute to abnormal cell proliferation by an inability to escape the cell cycle. Molecularly, It has been proposed that proteasome inhibition by MG132 can stabilize Runx2 protein levels in late G1 and S in MC3T3 cells, but not in osteosarcoma cells which consequently leads to a cancerous phenotype.[6][5]
Regulation and co-factors
Due to its role as a master transcription factor of osteoblast differentiation, the regulation of Runx2 is intricately connected to other processes within the cell.
Twist, Msh homeobox 2 (Msx2), and promyeloctic leukemia zinc-finger protein (PLZF) act upstream of Runx2. Osterix (Osx) acts downstream of Runx2 and serves as a marker for normal osteoblast differentiation. Zinc finger protein 521 (ZFP521) and activating transcription factor 4 (ATF4) are cofactors of Runx2.[12] Binding of the transcriptional coregulator, WWTR1 (TAZ) to Runx2 promotes transcription.
Furthermore, in proliferating chondrocytes, Runx2 is inhibited by CyclinD1/CDK4 as part of the cell cycle.[13]
Interactions
RUNX2 has been shown to interact with:
miR-133 and CyclinD1/CDK4 directly inhibits Runx2.[23][13]
References
- GRCh38: Ensembl release 89: ENSG00000124813 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000039153 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- San Martin IA, Varela N, Gaete M, Villegas K, Osorio M, Tapia JC, Antonelli M, Mancilla EE, Pereira BP, Nathan SS, Lian JB, Stein JL, Stein GS, van Wijnen AJ, Galindo M (December 2009). "Impaired cell cycle regulation of the osteoblast-related heterodimeric transcription factor Runx2-Cbfbeta in osteosarcoma cells". Journal of Cellular Physiology. 221 (3): 560–71. doi:10.1002/jcp.21894. PMC 3066433. PMID 19739101.
- Lucero CM, Vega OA, Osorio MM, Tapia JC, Antonelli M, Stein GS, van Wijnen AJ, Galindo MA (April 2013). "The cancer-related transcription factor Runx2 modulates cell proliferation in human osteosarcoma cell lines". Journal of Cellular Physiology. 228 (4): 714–23. doi:10.1002/jcp.24218. PMC 3593672. PMID 22949168.
- Wysokinski D, Pawlowska E, Blasiak J (May 2015). "RUNX2: A Master Bone Growth Regulator That May Be Involved in the DNA Damage Response". DNA and Cell Biology. 34 (5): 305–15. doi:10.1089/dna.2014.2688. PMID 25555110.
- Galindo M, Pratap J, Young DW, Hovhannisyan H, Im HJ, Choi JY, Lian JB, Stein JL, Stein GS, van Wijnen AJ (May 2005). "The bone-specific expression of Runx2 oscillates during the cell cycle to support a G1-related antiproliferative function in osteoblasts". The Journal of Biological Chemistry. 280 (21): 20274–85. doi:10.1074/jbc.M413665200. PMC 2895256. PMID 15781466.
- Pratap J, Galindo M, Zaidi SK, Vradii D, Bhat BM, Robinson JA, Choi JY, Komori T, Stein JL, Lian JB, Stein GS, van Wijnen AJ (September 2003). "Cell growth regulatory role of Runx2 during proliferative expansion of preosteoblasts". Cancer Research. 63 (17): 5357–62. PMID 14500368.
- Qiao M, Shapiro P, Fosbrink M, Rus H, Kumar R, Passaniti A (March 2006). "Cell cycle-dependent phosphorylation of the RUNX2 transcription factor by cdc2 regulates endothelial cell proliferation". The Journal of Biological Chemistry. 281 (11): 7118–28. doi:10.1074/jbc.M508162200. PMID 16407259.
- Lou Y, Javed A, Hussain S, Colby J, Frederick D, Pratap J, Xie R, Gaur T, van Wijnen AJ, Jones SN, Stein GS, Lian JB, Stein JL (February 2009). "A Runx2 threshold for the cleidocranial dysplasia phenotype". Human Molecular Genetics. 18 (3): 556–68. doi:10.1093/hmg/ddn383. PMC 2638795. PMID 19028669.
- Jinkins, J. R. (1987). "Large volume full columnar lumbar myelography". Neuroradiology. 29 (4): 371–373. doi:10.1007/bf00348917. PMID 3627420. S2CID 20760228.
- Berti M, Buso G, Colautti P, Moschini G, Stlevano BM, Tregnaghi C (August 1977). "Determination of selenium in blood serum by proton-induced X-ray emission". Analytical Chemistry. 49 (9): 1313–5. doi:10.1021/ac50017a008. PMID 883617.
- Baniwal SK, Khalid O, Sir D, Buchanan G, Coetzee GA, Frenkel B (August 2009). "Repression of Runx2 by androgen receptor (AR) in osteoblasts and prostate cancer cells: AR binds Runx2 and abrogates its recruitment to DNA". Molecular Endocrinology. 23 (8): 1203–14. doi:10.1210/me.2008-0470. PMC 2718746. PMID 19389811.
- Khalid O, Baniwal SK, Purcell DJ, Leclerc N, Gabet Y, Stallcup MR, Coetzee GA, Frenkel B (December 2008). "Modulation of Runx2 activity by estrogen receptor-alpha: implications for osteoporosis and breast cancer". Endocrinology. 149 (12): 5984–95. doi:10.1210/en.2008-0680. PMC 2613062. PMID 18755791.
- Hess J, Porte D, Munz C, Angel P (June 2001). "AP-1 and Cbfa/runt physically interact and regulate parathyroid hormone-dependent MMP13 expression in osteoblasts through a new osteoblast-specific element 2/AP-1 composite element". The Journal of Biological Chemistry. 276 (23): 20029–38. doi:10.1074/jbc.M010601200. PMID 11274169.
- D'Alonzo RC, Selvamurugan N, Karsenty G, Partridge NC (January 2002). "Physical interaction of the activator protein-1 factors c-Fos and c-Jun with Cbfa1 for collagenase-3 promoter activation". The Journal of Biological Chemistry. 277 (1): 816–22. doi:10.1074/jbc.M107082200. PMID 11641401.
- Schroeder TM, Kahler RA, Li X, Westendorf JJ (October 2004). "Histone deacetylase 3 interacts with runx2 to repress the osteocalcin promoter and regulate osteoblast differentiation". The Journal of Biological Chemistry. 279 (40): 41998–2007. doi:10.1074/jbc.M403702200. PMID 15292260.
- Pelletier N, Champagne N, Stifani S, Yang XJ (April 2002). "MOZ and MORF histone acetyltransferases interact with the Runt-domain transcription factor Runx2". Oncogene. 21 (17): 2729–40. doi:10.1038/sj.onc.1205367. PMID 11965546.
- Zhang YW, Yasui N, Ito K, Huang G, Fujii M, Hanai J, Nogami H, Ochi T, Miyazono K, Ito Y (September 2000). "A RUNX2/PEBP2alpha A/CBFA1 mutation displaying impaired transactivation and Smad interaction in cleidocranial dysplasia". Proceedings of the National Academy of Sciences of the United States of America. 97 (19): 10549–54. Bibcode:2000PNAS...9710549Z. doi:10.1073/pnas.180309597. PMC 27062. PMID 10962029.
- Hanai J, Chen LF, Kanno T, Ohtani-Fujita N, Kim WY, Guo WH, Imamura T, Ishidou Y, Fukuchi M, Shi MJ, Stavnezer J, Kawabata M, Miyazono K, Ito Y (October 1999). "Interaction and functional cooperation of PEBP2/CBF with Smads. Synergistic induction of the immunoglobulin germline Calpha promoter". The Journal of Biological Chemistry. 274 (44): 31577–82. doi:10.1074/jbc.274.44.31577. PMID 10531362.
- Li X, Huang M, Zheng H, Wang Y, Ren F, Shang Y, Zhai Y, Irwin DM, Shi Y, Chen D, Chang Z (June 2008). "CHIP promotes Runx2 degradation and negatively regulates osteoblast differentiation". The Journal of Cell Biology. 181 (6): 959–72. doi:10.1083/jcb.200711044. PMC 2426947. PMID 18541707.
- Li Z, Hassan MQ, Volinia S, van Wijnen AJ, Stein JL, Croce CM, Lian JB, Stein GS (September 2008). "A microRNA signature for a BMP2-induced osteoblast lineage commitment program". Proceedings of the National Academy of Sciences of the United States of America. 105 (37): 13906–11. Bibcode:2008PNAS..10513906L. doi:10.1073/pnas.0804438105. PMC 2544552. PMID 18784367.
Further reading
- Otto F, Kanegane H, Mundlos S (March 2002). "Mutations in the RUNX2 gene in patients with cleidocranial dysplasia". Human Mutation. 19 (3): 209–16. doi:10.1002/humu.10043. PMID 11857736. S2CID 2578591.
- Komori T (March 2002). "[Cbfa1/Runx2, an essential transcription factor for the regulation of osteoblast differentiation]". Nihon Rinsho. Japanese Journal of Clinical Medicine. 60 Suppl 3: 91–7. PMID 11979975.
- Stock M, Otto F (June 2005). "Control of RUNX2 isoform expression: the role of promoters and enhancers". Journal of Cellular Biochemistry. 95 (3): 506–17. doi:10.1002/jcb.20471. PMID 15838892. S2CID 29657025.
- Blyth K, Cameron ER, Neil JC (May 2005). "The RUNX genes: gain or loss of function in cancer". Nature Reviews. Cancer. 5 (5): 376–87. doi:10.1038/nrc1607. PMID 15864279. S2CID 335980.
- Schroeder TM, Jensen ED, Westendorf JJ (September 2005). "Runx2: a master organizer of gene transcription in developing and maturing osteoblasts". Birth Defects Research. Part C, Embryo Today. 75 (3): 213–25. doi:10.1002/bdrc.20043. PMID 16187316.
- Frenkel B, Hong A, Baniwal SK, Coetzee GA, Ohlsson C, Khalid O, Gabet Y (August 2010). "Regulation of adult bone turnover by sex steroids". Journal of Cellular Physiology. 224 (2): 305–10. doi:10.1002/jcp.22159. PMC 5770230. PMID 20432458.
External links
- GeneReviews/NCBI/NIH/UW entry on Cleidocranial Dysplasia
- Runx2+protein at the U.S. National Library of Medicine Medical Subject Headings (MeSH)
PDB gallery | |
|---|---|
|