Coccolithovirus
Coccolithovirus is a genus of giant double-stranded DNA virus, in the family Phycodnaviridae. Algae, specifically Emiliania huxleyi, a species of coccolithophore, serve as natural hosts. There is only one described species in this genus: Emiliania huxleyi virus 86.[1][2]
| Coccolithovirus | |
|---|---|
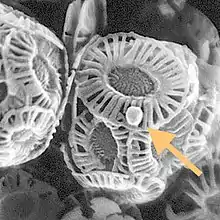 | |
| Giant coccolithovirus, Emiliania huxleyi virus 86 (arrowed), infecting an Emiliania huxleyi coccolithophore | |
| Virus classification | |
| (unranked): | Virus |
| Realm: | Varidnaviria |
| Kingdom: | Bamfordvirae |
| Phylum: | Nucleocytoviricota |
| Class: | Megaviricetes |
| Order: | Algavirales |
| Family: | Phycodnaviridae |
| Genus: | Coccolithovirus |
| Strains | |
| |
Structure
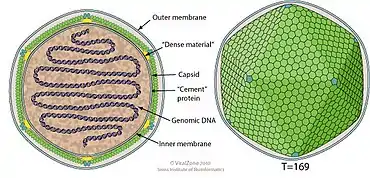
Coccolithoviruses are enveloped, icosahedral and have a diameter ranging from 100–220 nm. Their genomes are linear, between 410–415kb in length and predict to encode for approximately 472 proteins.[1]
| Genus | Structure | Symmetry | Capsid | Genomic arrangement | Genomic segmentation |
|---|---|---|---|---|---|
| Coccolithovirus | Icosahedral | T=169 | Enveloped | Linear | Monopartite |
Life cycle
Coccolithoviruses are part of the family of Phycodnaviridae, one of the five families that belong to a large and phylogenetically diverse group of viruses known as nucleocytoplasmic large dsDNA viruses (NCLDVs). These viruses either replicate exclusively in the cytoplasm of the host cell or start their life cycle in the host nucleus but complete it in the cytoplasm. In the case of EhV-86 the infection strategy is not fully understood but Mackinder et al. (2009)[3] have proposed the following model: The virus enters the host cell via endocytosis, followed by fusion of its lipid membrane with the host vacuole membrane and the release of its nucleoprotein core into the cytoplasm. Alternatively the virus membrane could fuse directly with the host plasma membrane. The virus genome is then released from the capsid into the nucleus, where it is replicated by the viral DNA polymerase. The replicated genome is packed into assembled capsids in the cytoplasm and the newly formed (up to 400–1000) virions are thought to be transported to the plasma membrane and released by a controlled budding mechanism, which leads to the cellular breakdown of the host cell.
During G2 and M stage of the life cycle, the coccosphere is incomplete and the exposure of the plasma membrane to the virus is increased. Even with an intact coccosphere infection can occur due to naturally occurring gaps between the coccoliths.[4]
E. huxleyi is known for forming seasonal algal blooms, that can reach 250,000 km2, during which cell density in the upper 200 m increases from 103 to 105 cells per mL seawater.[5] These algae blooms collapse usually after 5–8 days and several studies have shown that bloom termination is intrinsically linked to infection by coccolithoviruses.[6] Transmission of viruses between algal hosts occurs via passive diffusion. Furthermore, EhV DNA was also detected in copepods, leading to the proposal that viruses are further dispersed by virus-carrying zooplankton.[7]
Genome
To date 14 EhV strains have been isolated between 1999 and 2008 primarily from the English Channel (EC) but also from the Norwegian and Scotland's Coast.[8][9][10][11][12] Although partial sequences of all these 14 strains are available, EhV-86 is the only strain that has been fully sequenced due to the highly repetitive nature of the genome.[13] The sequencing of EhV-86 revealed a circular genome of 407,339 bp length with 472 predicted coding sequences (CDS). Remarkably 80% of these putative genes have no database homologs to date. Those that could be assigned a function due to sequence similarity or protein domain matches include DNA and RNA polymerase subunits, eight proteases as well as at least four genes that encode proteins involved in sphingolipid biosynthesis. These were shown to have been acquired from the host via horizontal gene transfer.[14]
Furthermore, the EhV-86 genome revealed three distinct families (A, B, C) of repetitive regions within the genome.[15] Family C consists of AT-rich repeats that are non-coding and that are probably part of the origin of replication (ORF). Family B are GC-rich repeats that are found in protein products of eight predicted CDSs. Family A homologous regions vary in size between 30–300 bp and are found in a 104 kbp (200–304 kbp) section of the genome, that contains no gene homologs of known function in the current data bases. Family A repeat units are non-coding and characterised by a nanomer (GTTCCC(T/C)AA) that in total, appears at 106 locations within this region. This sequence is found directly upstream of 86 CDSs and is likely to play a role in controlling the expression of associated CDSs.
During infection a distinct expression pattern of viral genes was described that could be divided into three phases according to CDS expression.[16] One hour post infection, 39 viral genes were transcribed, followed by a further 194 genes after 2 h and 71 genes after 4 h. All 39 genes that are expressed 1 h post infection are located in the 104 kbp region and have the nanomer directly upstream of the start codon. Since the expression of viral RNA polymerase was not detected 1 h after infection, it has yet to be established whether the promoter is recognised by a packaged viral RNA polymerase or by the host RNA polymerase. Proteomic analysis of the EhV-86 virion did, however, fail to detect any major RNA polymerase subunits.[17]
History
Wilson and his team at the Marine Biological Association (MBA), University of East Anglia and Plymouth Marine Laboratory (PML), first observed the virus in 1999. Later in the summer of 2005 researchers at the Plymouth Marine Laboratory (Willie Wilson et al.) and at the Sanger Institute (Holden et al.) sequenced the genome for the EhV-86 strain finding it to have 472 protein-coding genes making it a "giant-virus", and the largest known marine virus by genome.
From initial investigation of the Coccolithoviruses genome, a sequence of genes responsible for production of ceramide was discovered.[18] Ceramide is a controlling factor in cell death, and it is currently thought that Coccolithovirus uses this to prolong the life of Emiliania huxleyi while it uses the host cell to replicate. This is a unique ability unseen in any other viral genome to date.
See also
- Mimivirus – largest giant virus on record by genome
- Mycoplasma genitalium, Pelagibacter ubique – some of the smallest known bacteria
- Nanoarchaeum – smallest known archaeum
- Smallest organisms
- Parvovirus – smallest known family of viruses
- Phycodnaviridae – algae infecting viruses
Notes
References
- "Viral Zone". ExPASy. Retrieved 15 June 2015.
- "Virus Taxonomy: 2020 Release". International Committee on Taxonomy of Viruses (ICTV). March 2021. Retrieved 22 May 2021.
- Mackinder LCM, Worthy CA, Biggi G, Hall M, Ryan KP, Varsani A, Harper GM, Wilson WH, Brownlee C, Schroeder DC. (2009) A unicellular algal virus, Emilienia huxleyi virus 86, exploits an animal-like infection strategy. Journal of general Virology 90:2306–2316.
- Paasche E. (2001) A review of the coccolithophorid Emiliania huxleyi (Prymnesiophyceae), with particular reference to growth, coccolith formation, and calcification-photosynthesis interactions. Phycologia 40(6):503–52.
- Schroeder DC, Oke J, Hall M, Malin G, Wilson WH. (2003) Viral succession observed during an Emilinaia huxleyi bloom. Applied and Environmental Microbiology 69:2484–2490.
- Wilson WH, Tarran GA, Schroeder D, Cox M, Oke J, Malin G. (2002) Isolation of viruses responsible for the demise of an Emiliania huxleyi bloom in the English Channel. Journal of the Marine Biological Association of the United Kingdom 82:369–377.
- Frada MJ, Schatz D, Farstey V, Ossolinski JE, Sabanay H, Ben-Dor S, Koren I, Vardi A. (2014) Zooplankton May Serve as Transmission Vectors for Viruses Infecting Algal Blooms in the Ocean. Current Biology 24:2592–2597.
- Nissimov JI, Napierb JA, Kimmance SA, Allen MJ. (2014) Permanent draft genomes of four new coccolithoviruses: EhV-18, EhV-145, EhV-156 and EhV-164. Marine Genomics 15:7–8.
- Nissimov JI, Worthy CA, Rooks P, Napier JA, Kimmance SA, Henn MR, Ogata H, Allen MJ. (2011) Draft genome sequence of the Coccolithovirus EhV-84. Standards in Genomic Science 5:1–11.
- Nissimov JI, Worthy CA, Rooks P, Napier JA, Kimmance SA, Henn MR, Ogata H, Allen MJ. (2012) Draft Genome Sequence of Four Coccolithopviruses: Emiliania huxleyi Virus EhV-88, EhV-201, EhV-207 and EhV-208. Journal of Virology 86(5):2896–2897.
- Pagarete et al. 2012, Allen et al. 2006d, Nissimov JI, Worthy CA, Rooks P, Napier JA, Kimmance SA, Henn MR, Ogata H, Allen MJ. (2012) Draft Genome Sequence of the Coccolithopvirus Emiliania huxleyi Virus 202. Journal of Virology 86(4):380–2381.
- Nissimov JI, Worthy CA, Rooks P, Napier JA, Kimmance SA, Henn MR, Ogata H, Allen MJ. (2011) Draft Genomic Sequence of the Coccolithovirus Emiliania huxleyi Virus 203. Journal of Virology 85(24):13468–13469.
- Wilson WH, Schroeder DC, Allen MJ, Holden MTG, Parkhill J, Barrell BG, Churcher C, Hamlin N, Mungall K, Norbertczak H, Quail MA, Price C, Rabbinowitsch E, Walker D, Craigon M, Roy D, Ghazal P. (2005) Complete Genome Sequence and Lytic Phase Transcription Profile of a Coccolithovirus. Science 309:1090–1092
- Monier A, Pagarete A, De Vargas C, Allen MJ, Read B, Claverie J, Ogata H, De Vargas C. (2009) Horizontal gene transfer of an entire metabolic pathway between a eukaryotic alga and its DNA virus. Genome Research 19:1441–1449.
- Allen MJ, Schroeder DC, Wilson WH. (2006) Preliminary characterisation of repeat families in the genome of EhV-86, a giant algal virus that infects the marine microalga Emiliania huxleyi. Archives of Virology 151:525–535.
- Allen MJ, Forster T, Schroeder DC, Hall M, Roy D, Ghazal P, Wilson WH. (2006) Locus specific gene expression pattern suggests a unique propagation strategy for a giant algal virus. Journal of Virology 80: 7699–7705.
- Allen MJ, Howard JA, Lilley KS, Wilson WH. (2008) Proteomic analysis of the EhV-86 virion. Proteome Science 6(11).
- Wilson WH, Schroeder DC, Allen MJ, Holden MTG, Parkhill J, Barrell BG, Churcher C, Hamlin N, Mungall K, Norbertczak H, Quail MA, Price C, Rabbinowitsch E, Walker D, Craigon M, Roy D, Ghazal P. (2005) Complete Genome Sequence and Lytic Phase Transcription Profile of a Coccolithovirus. Science 309:1090–1092
Further reading
- Wilson, W. H., Schroeder, D. C., Allen, M. J., Holden, M. T. G., Parkhill, J., Barrell, B. G., Churcher, C., Hamlin, N., Mungall, K., Norbertczak, H., Quail, M. A., Price, C., Rabbinowitsch, E., Walker, D., Craigon, M., Roy, D. and Ghazal, P. (2005) Complete Genome Sequence and Lytic Phase Transcription Profile of a Coccolithovirus. Science 5737, 1090–1092
- Allen, M. J., Schroeder, D. C., Holden, M. T. and Wilson, W. H. (2006) Evolutionary History of the Coccolithoviridae. Mol Biol Evol. 23, 86–92 (Athens login required)