Fermentation alcoolique
La fermentation alcoolique est un processus biochimique par lequel des sucres (glucides, principalement le glucose) sont transformés en alcool (éthanol) dans un milieu liquide, privé d'air.

La réaction libère de l'énergie. La plupart des ferments microorganiques (levures) qui ont la capacité de fermentation alcoolique utilisent cette réaction pour gagner temporairement de l'énergie quand l'oxygène nécessaire à la respiration cellulaire manque.
Histoire de la découverte

L'homme a utilisé la fermentation alcoolique par exemple dans la fabrication de la bière ou celle du vin depuis des millénaires, sans connaître les processus biologiques précis.
En 1815, le chimiste français Joseph Louis Gay-Lussac a établi pour la première fois l'équation brute de la réaction chimique de la décomposition du glucose en éthanol. Puis se développèrent diverses conceptions sur le mécanisme de la fermentation. Tandis que dans les années 1830 Jöns Jakob Berzelius et Justus von Liebig attribuaient, dans une « théorie mécaniste de la fermentation, » une action catalytique à certaines substances, Charles Cagniard de Latour, Theodor Schwann et Friedrich Traugott Kützing pouvaient prouver, indépendamment les uns des autres, que des êtres vivants, notamment des levures, en étaient responsables[1]. Louis Pasteur a aussi postulé en 1857 une « théorie vitaliste de la fermentation » selon laquelle la fermentation alcoolique n'était possible qu'en liaison avec des cellules vivantes.
En 1871, Marie von Manasseïnn parvint à faire publier un article sur ses travaux réalisés l'année précédente dans le laboratoire de Julius Wiesner à Vienne[2]. Elle fut la première à conclure qu'il n'est pas nécessaire d'avoir de la levure vivante pour obtenir une fermentation alcoolique[3].
Le , Eduard Buchner publia des preuves de fermentation alcoolique au moyen d'extraits de levure sans cellules. Il rendit la matière zymase — identifiée maintenant comme un mélange de diverses enzymes — responsable de la transformation du sucre en éthanol, et reçut en 1907 le prix Nobel de chimie « pour ses recherches en biochimie et la découverte de la fermentation en l'absence de cellules ». Marie von Manasseïnn revendiqua l'antériorité de ces conclusions, mais Eduard Buchner rejeta les travaux comme invalides. Pour lui, les conclusions de Marie von Manasseïnn relevaient d'un présupposé subjectif ; il contesta le caractère probant de ses expériences, aussi bien en raison de la haute température appliquée pour obtenir des extraits de levure, que du temps trop court de stérilisation de la solution sucrée utilisée, conduisant à un risque de contamination[3].
Des recherches plus approfondies de Arthur Harden et William John Young (en) conduisirent à la découverte d'un produit intermédiaire phosphorylé : l'ester de Harden-Young, aujourd'hui connu sous le nom de fructose-1,6-bisphosphate. Harden et Hans von Euler-Chelpin reçoivent ensemble en 1929 aussi le prix Nobel de chimie « pour leurs travaux sur la fermentation des sucres et les enzymes qui y participent. » Après que les réactions partielles ont été mises en évidence et que des schémas pour le déroulement de la fermentation ont été esquissés, Otto Heinrich Warburg a identifié la coenzyme Nicotinamide adénine dinucléotide (NADH) comme composant essentiel du processus de fermentation. Dès 1937, Erwin Negelein (de) et Hans Joachim Wulff réussissaient la cristallisation de l'enzyme de fermentation alcool déshydrogénase[4].
Aujourd'hui, les enzymes d'espèces diverses prenant part à la fermentation ont été isolées et caractérisées biochimiquement (optimum de pH, de température, vitesse de réaction, taux de transformation). L'analyse de la structure cristalline a donné une première vision sur leur structure spatiale moléculaire. On a des connaissances sur les mécanismes de réaction. Au total, on est ainsi dans la position de faire des comparaisons entre espèces[5],[6]. Les gènes décodés, qui contiennent la structure de ces enzymes donnent des conclusions sur leur origine évolutionnaire et leur fonction originelle éventuelle[réf. souhaitée].
Le rôle dans le métabolisme
La fermentation alcoolique est principalement utilisée par diverses espèces de levures pour produire de l'énergie. Si elles ont de l'oxygène à leur disposition, elles oxydent les sucres par respiration cellulaire, et trouvent ainsi l'énergie nécessaire à la vie. Les sucres sont alors complètement oxydés par une longue chaîne de réactions enzymatiques (glycolyse - décarboxylation du pyruvate - cycle de Krebs - chaîne respiratoire) en dioxyde de carbone et en eau, en consommant de l'oxygène. S'il n'y a pas d'oxygène disponible, les levures ont dans la fermentation alcoolique une autre possibilité de fourniture d'énergie. Mais elles peuvent ainsi — par comparaison avec la respiration cellulaire — récupérer substantiellement moins d'énergie du glucose, sous forme d'adénosine triphosphate (ATP) : par oxydation complète, une molécule de glucose fournit 36 molécules d'ATP[7], mais par fermentation alcoolique seulement 2 molécules d'ATP. Ces deux molécules sont obtenues dans la glycolyse, la première étape de la chaîne de réactions aussi bien pour la respiration cellulaire que pour la fermentation. Les deux étapes supplémentaires de la fermentation, et donc la production d'éthanol servent non pas à faire de l'énergie, mais à la régénération du cofacteur NAD+ utilisé par les enzymes de la glycolyse. Comme le NAD+ est disponible en quantité limitée, il est transformé par les enzymes de fermentation de l'état réduit NADH en état oxydé NAD+ par réduction de l'acétaldéhyde en éthanol.
Les levures sont donc des aérobies facultatifs. Quand l'oxygène est disponible, le glucose est métabolisé par voie aérobie. En l'absence d'air, les levures doivent par contre faire la fermentation alcoolique. Comme celle-ci produit bien moins d'énergie que la respiration aérobie, le besoin en glucose augmente considérablement. Ce phénomène est nommé effet Pasteur. En raison de la production d'énergie limitée, les levures se multiplient en l'absence d'air bien moins vite qu'en sa présence. En plus, l'éthanol fabriqué joue le rôle de poison cellulaire.
On a aussi observé la production d'éthanol par des levures, malgré la présence de suffisamment d'oxygène. Ceci se produit quand elles vivent dans un milieu sursucré, et que les enzymes de la respiration cellulaire sont surchargées. Les levures consomment constamment le sucre, et le transforment par fermentation à côté de la respiration. Il s'agit ici de l'Effet Crabtree[8].
Outre les levures, beaucoup de bactéries pratiquent la fermentation alcoolique[9]. C'est ainsi que Sarcina ventriculi utilise la même voie enzymatique que la levure, tandis que Zymomonas mobilis emprunte une autre voie. De même, on a pu démontrer dans certaines plantes de faibles productions d'éthanol sous l'action du manque d'oxygène[10].
Bases biochimiques
Réactions enzymatiques

Les premières étapes de la fermentation alcoolique sont celles de la glycolyse. Chez la levure de boulanger (Saccharomyces cerevisiae), la voie suivie est celle de Embden-Meyerhof-Parnas, tandis que chez le Zymomonas mobilis, une bactérie, c'est celle de Entner-Doudoroff[11]. Cette voie transforme une molécule de D-glucose en deux molécules de pyruvate. Le S. cerevisiae fabrique deux molécules d'adénosine triphosphate (ATP) à partir de deux molécules d'adénosine diphosphate (ADP) et de deux radicaux phosphate (Pi) par phosphorylation. Chez Z. mobilis, une seule molécule d'ATP est formée. De plus, dans les deux voies, deux ions NAD+ sont réduits en 2 molécules de NADH.
Pour que la glycolyse puisse continuer à se faire, il faut que le NAD+ soit régénéré. Ceci se produit en conditions anaérobies par la réaction de fermentation qui suit. De chaque molécule de pyruvate, une molécule de dioxyde de carbone est détachée par l'enzyme décarboxylase du pyruvate. Pour cette réaction, il faut comme coenzymes la thiamine pyrophosphate, un voisin de la vitamine B1 (ou thiamine) et deux ions magnésium. Ne pas confondre la décarboxylase du pyruvate avec le complexe pyruvate déshydrogénase, qui joue un rôle central dans l'oxydation aérobie du pyruvate.
L'acétaldéhyde produit lors de cette étape est très toxique pour l'organisme, et est immédiatement transformé lors de l'étape suivante. L'enzyme de catalyse alcool déshydrogénase (EC 1.1.1.1) contient un ion zinc (Zn2+) qui polarise le groupe carbonyle de l'acétaldéhyde. Ceci permet le transfert de deux électrons et d'un proton du NADH vers l'acétaldéhyde, ce qui le réduit en éthanol, et le NAD+ est régénéré. La glycolyse et les deux réactions suivantes prennent place dans le cytoplasme de la cellule.
Pour la levure de boulanger, l'équation globale de la réaction s'écrit :
| Glucose, | 2 ADP et | 2 phosphate | donnent | 2 éthanol, | 2 dioxyde de carbone, | et 2 ATP | . |







L'enzyme alcool déshydrogénase fait de l'éthanol par réduction de l'acétaldéhyde, mais peut aussi catalyser la réaction inverse. Pendant la fermentation alcoolique, le processus de réduction de l'acétaldéhyde en éthanol est majoritaire. L'éthanol produit est alors excrété des cellules vers le milieu environnant.
L'oxydation de l'éthanol en acétaldéhyde a lieu par contre par exemple dans la détoxication de l'éthanol dans le foie. L'acétaldéhyde est toxique, et constitue, à côté des alcools de fusel, la principale cause du mal de tête et de la nausée après une ingestion importante d'alcool (la fameuse « gueule de bois »). L'acétaldéhyde est oxydé en acide acétique par l'enzyme acétaldéhyde déshydrogénase.
Conditions de la fermentation
La fermentation alcoolique ne peut se produire que sous certaines conditions :
- Température :
- trop froid (10 °C), le processus est très ralenti voire incomplet
- trop chaud (au-delà de 45 à 50 °C), les levures meurent, le milieu devient inapte à leur survie. La formation d'alcool pendant la fermentation limite encore plus fortement la capacité de résistance à la température de Saccharomyces cerevisiae. On considère, en vinification, que 30 °C - 32 °C est une température maximale, au-delà de laquelle le risque d'arrêt de la fermentation est très élevé. De plus en plus de vinificateurs travaillent d'ailleurs, en vin rouge, à des températures proches de 25 °C grâce à la maîtrise permise par l'installation d'échangeurs thermiques immergés (appelés couramment « drapeaux ») dans les cuves de vinification
- Oxygène : Bien que la fermentation soit un phénomène anaérobie, les levures ont besoin d'un peu d'oxygène pour se multiplier et synthétiser des stérols qui permettent une meilleure résistance à l'éthanol (et donc une survie améliorée).
- Azote assimilable : Il provient des raisins ou de leur moût mis à fermenter. Il est nécessaire pour assurer la production de protéines, donc la multiplication des levures. Dans les zones les plus méridionales, il est souvent en quantité insuffisante par rapport aux besoins de la population de levures : la carence absolue est fixée autour de 150 mg/ℓ (pour fermenter 200 g de sucres /ℓ) avec des besoins qui croissent en fonction de la quantité de sucres à fermenter. Une carence peut conduire à plusieurs résultats souvent négatifs pour la qualité des vins obtenus : production de composés soufrés malodorants (odeurs soufrées, « de réduit », de chou...), ralentissement des fins de fermentation ou arrêt de fermentation avec présence de sucres résiduels. Ces deux dernières situations induisent une élévation des risques de multiplication d'espèces indésirées à ce stade de la vinification : bactéries lactiques, bactéries acétiques, levures contaminantes (Brettanomyces bruxellensis principalement).
- Alcool : C'est un antiseptique, même pour les micro-organismes qui l'ont produit. Passée une certaine concentration (de 14 à 16 %), il agit comme un poison pour les levures. Les boissons de plus haut degré alcoolique sont produites par distillation.
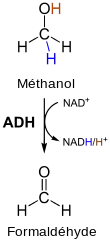
Dans la fermentation alcoolique par les levures, des produits annexes indésirables sont également produits, comme le méthanol, et autres alcools comme le 1-butanol, l'alcool amylique et des hexanols. Leur formation ne suit pas la voie métabolique décrite ici, mais ils viennent par exemple de la décomposition d'amino-acides. Dans le corps, le méthanol est oxydé par l'enzyme alcool déshydrogénase en formol très toxique. Si l'on boit beaucoup d'alcool de mauvaise qualité (avec une grande proportion de méthanol), il se produit dans le corps une grande quantité de formol, qui endommage des protéines très sensibles, notamment les senseurs de l'œil, et il peut se produire dans le pire des cas des crampes musculaires, la cécité et finalement la mort.
Régulation
La régulation, c'est-à-dire le changement entre respiration cellulaire aérobie et fermentation anaérobie est un thème de recherches actuel. On ne peut pas établir de schéma de régulation selon le système « Changer l'interrupteur quand il manque X. » Il y a déjà des différences entre diverses variétés de levures, sans parler des plantes et des bactéries. Mais les chercheurs sont près de trouver la solution du problème[12]. Un rôle essentiel est joué par les concentrations en oxygène et en glucose.
En outre, il y a par exemple chez S. cerevisiae deux gènes pour l'enzyme alcool déshydrogénase cytosolique, et donc deux enzymes légèrement différentes, ADH1 et ADH2. Les deux enzymes peuvent transformer l'acétaldéhyde en éthanol et réciproquement. Mais de petites différences dans leur structure moléculaire donnent à ces réactions des vitesses différentes : ADH1 transforme l'acétaldéhyde en éthanol plus vite, tandis qu'ADH2 effectue plus vite la réaction inverse. La présence des enzymes est régulée par des facteurs de transcription, qui dirigent la lecture des gènes[13]. L'ADH1 est toujours présente. Si le niveau de glucose tombe de façon drastique, alors s'enclenche la synthèse de l'enzyme ADH2, qui peut métaboliser l'éthanol pour faire de l'énergie (en présence d'oxygène), et maintient ainsi la levure en vie. La levure peut aussi faire de l'éthanol quand suffisamment de sucre est présent, et décomposer cet éthanol plus tard quand elle a un besoin urgent d'énergie. Du point de vue de l'évolution, elle a ainsi un avantage : elle empoisonne tous les concurrents à la nourriture avec de l'éthanol, puis utilise ce dernier elle-même. La naissance des deux gènes ADH1 et ADH2 est probablement due à la duplication d'un même gène origine. Chez d'autres espèces, il existe même plus de deux alcool déshydrogénases.
Bilan énergétique
Comme nous l'avons vu, sous conditions anoxiques, la respiration cellulaire avec la phosphorylation d'ADP en ATP de la chaîne respiratoire ne fonctionne pas, la seule source d'énergie dans ces conditions est la glycolyse avec formation d'ATP par phosphorylation du substrat. Elle ne fournit que 2 molécules d'ATP par molécule de glucose, tandis que la respiration cellulaire en fournit 38.
Si la décomposition du glucose en pyruvate venait à s'arrêter, le processus s'arrêterait vite, parce que la glycolyse utiliserait tout le NAD+ disponible. Cette substance n'existe qu'en traces dans la cellule, et doit être constamment régénérée. C'est pourquoi dans la fermentation alcoolique, le pyruvate est décarboxylé et l'acétaldéhyde est réduit en éthanol par le NADH, avec oxydation du NADH en NAD+. En prenant la chaîne de réaction globale du glucose à l'éthanol, il ne se produit pas de NADH, riche en énergie.
Si l'on considère le carbone, son nombre d'oxydation passe de 0 dans le glucose à +4 dans une molécule de dioxyde de carbone et à -2 dans chacune des 2 molécules d'éthanol. La fermentation alcoolique constitue ainsi une dismutation, un cas particulier des réactions d'oxydoréduction.
La variation de l'enthalpie libre dans la fermentation alcoolique s'élève sous les conditions standard, sauf à pH 7 au lieu de 0, à ΔG0' = - 218 kJ par mole de glucose, et en cas de respiration cellulaire à - 2822 kJ par mole de glucose. Les conditions standard sont : température 25 °C, pression 1,013 bar, concentration des matières participant à la réaction 1 mol/ℓ, à l'exception de l'eau, pour laquelle on prend 55,6 mol/ℓ (eau pure), et pour les gaz, pour lesquels la concentration correspond à l'équilibre avec une pression partielle de 1 bar dans la phase gazeuse. Pour les systèmes biologiques, la concentration des ions H+ ne peut être 1 mol/ℓ correspondant au pH 0, non tolérée par les êtres vivants, mais 10−7 mol/ℓ correspondant au pH 7. Si les conditions réelles dévient de ces conditions standard, la variation d'enthalpie libre peut aussi dévier substantiellement de la valeur standard. Dans les systèmes vivants, les conditions ne sont pas standard en règle générale, et elles changent souvent pendant le métabolisme. La variation d'enthalpie libre sous les conditions standard n'offre donc chez les êtres vivants qu'un point de repère pour la quantité d'énergie libérée dans une chaîne métabolique.
Autres substrats
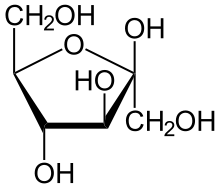
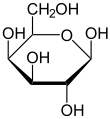
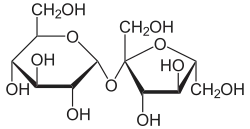
Outre le glucose, d'autres sucres simples peuvent être soumis à glycolyse, et donc subir la fermentation alcoolique. Cependant, la plupart des levures ont une affinité particulière pour le glucose (elles sont « glucophiles »), si bien que par exemple pendant la fermentation alcoolique du moût de raisin, qui contient en parties égales du glucose et du fructose, le glucose est transformé préférentiellement. Quand le vin terminé est doux, c'est-à-dire que tout le sucre n'est pas transformé en alcool, la plus grande partie du sucre restant est du fructose. Ceci est particulièrement intéressant pour les diabétiques.
Le D-fructose peut d'une part être phosphorylé par une hexokinase, la première enzyme de la glycolyse, comme le glucose, et être ainsi entraîné sur la voie de la glycolyse, mais d'autre part, il peut être transformé par l'enzyme fructokinase en fructose-1-phosphate, à son tour clivé par une aldolase en dihydroxyacétone-phosphate, ultérieurement isomérisé en glycéraldéhyde 3-phosphate, et du glycéraldéhyde. Ce dernier va être phosphorylé en glycéraldéhyde-3-phosphate. Le glycéraldéhyde-3-phosphate rejoint la voie principale de la glycolyse.
Le D-galactose est transformé par les étapes galactose-1-phosphate et uridine diphosphate galactose en glucose, qui rejoint la glycolyse.
À côté des sucres simples, les disaccharides peuvent aussi être utilisés, dans la mesure où les enzymes nécessaires à leur hydrolyse en sucres simples sont présentes. C'est ainsi que le saccharose est hydrolysé par l'invertase en ses composants : glucose et fructose, qui subissent la glycolyse, comme décrit ci-avant. De même, le lactose est hydrolysé par la lactase en glucose et galactose. Ceci est vrai également pour les polysaccharides. Par exemple, pour utiliser l'amidon des céréales, on peut amener les graines à germer. L'enzyme amylase produite alors par la plante hydrolyse l'amidon en maltose, qui peut être à son tour transformé par la levure.
Une autre voie
La bactérie Zymomonas mobilis est en mesure de produire de l'éthanol à partir du glucose. Cependant, elle n'utilise que la moitié de la voie métabolique décrite auparavant. Au lieu de glycolyse, le glucose est transformé en pyruvate et glycéraldéhyde-3-phosphate par la voie d'Entner-Doudoroff[14]. Le glycéraldéhyde-3-phosphate peut être introduit dans la glycolyse et aussi dégradé en pyruvate. Les deux dernières étapes de la fermentation alcoolique correspondent à celles des levures. Ceci conduit avec une molécule de glucose à une seule molécule d'ATP. Mais la fermentation se déroule plus vite avec cette voie qu'avec celle utilisée par les levures, et conduit à une plus grande consommation de glucose. Z. mobilis est utilisé pour la fabrication du pulque à partir du jus d'agave.
Sous-produits de la fermentation
Les sous-produits de la fermentation sont produits à côté de l'éthanol et du dioxyde de carbone pendant la fermentation. Certains de ces produits sont aussi désignés par alcools de fusel[15].
On peut même les mettre en évidence dans la fermentation d'une solution de glucose pur. Dans la brasserie, la différence de goût entre le moût et la bière montre que des sous-produits de fermentation ont été produits. Ils contiennent par exemple des alcools plus élevés, comme le propan-1-ol, l'isobutanol, l'alcool amylique chiral (2-méthylbutan-1-ol), le 3-méthylbutan-1-ol, ainsi que des alcools aromatiques comme le 2-phényléthanol, le tyrosol ou le tryptophol. En outre, on trouve des esters carboxyliques, comme l'acétate d'éthyle, l'acétate de phényle ou l'acétate de pentyle. Également, des composés carbonylés comme des aldéhydes, comme éthanal, propanal, butanal ou furfural, ainsi que des cétones ou doubles cétones.
Des composés soufrés, comme l'hydrogène sulfuré H2S, le dioxyde de soufre SO2, le mercaptan éthylique et le méthylmercaptan apparaissent en petites quantités.
En outre on trouve des acides organiques, comme l'acide acétique, l'acide lactique, l'acide pyruvique, l'acétolactate, et des acides gras (de C4 à C12).
Des alcools multiples comme la glycérine, le butane-2,3-diol et le pentane-2,3-diol peuvent se trouver aussi dans les sous-produits de la fermentation alcoolique.
Les matières énoncées plus haut ne sont que les plus importantes de chaque groupe.
Occurrence naturelle
On trouve partout dans la nature des micro-organismes. C'est ainsi que les fruits sont couverts de bactéries et de levures, que l'on ne peut pas complètement enlever par simple lavage. Si le fruit reste dans un environnement chaud après cueillette, les microbes se multiplient. Ils détruisent la structure des cellules et s'introduisent à l'intérieur du fruit colonisé. On le trouvera par exemple sur une pomme à un endroit plus mou ou une tache brune. Pendant le processus de destruction, il peut se produire, surtout à l'intérieur du fruit, un manque d'oxygène. Les levures présentes changent alors leur métabolisme en fermentation alcoolique. Il est donc possible que des fruits qui se gâtent contiennent de l'alcool.
Jusqu'au XXe siècle, on n'ajoutait pas de levure supplémentaire dans la fabrication du vin, on n'utilisait pas non plus l'ensemencement (qui était connu par ailleurs, étant pratiqué pour la fabrication de pain, fromage, vinaigre, bière, etc.). Jusqu'au milieu du XIXe siècle, on n'avait que le savoir empirique que les liquides sucrés commencent après quelque temps à former de l'alcool, sans aucune connaissance des processus biologiques et des microorganismes à l'œuvre. Pour la fabrication du vin, ceci est encore en partie laissé à la nature. Après le pressurage des raisins en moût, les levures situées initialement sur les peaux se distribuent dans le liquide et commencent une fermentation spontanée. Sans addition de levures sélectionnées, ceci dure un peu plus longtemps, car la concentration des levures naturelles est très faible au début, mais le vin acquiert une note plus individuelle. Les variétés de levures diffèrent selon le terroir des raisins, ce qui fait qu'un vin produit par ce procédé peut être classé au goût selon son terroir. Les espèces de levures couramment présentes sont Kloeckera apiculata et Saccharomyces exiguus. Si le vigneron fait confiance aux levures naturelles, il court évidemment le risque que d'autres levures ou bactéries vivant sur les peaux de raisin prennent le dessus pendant la fabrication, et gâtent le moût. C'est pourquoi diverses variétés de levures sont sélectionnées pour donner à chaque vin son arôme typique. Ces levures sélectionnées ne comprennent qu'une variété de levure, et sont souvent spécialisées pour une sorte de raisins. Depuis que l'on sait utiliser la lyophilisation pour la conservation des levures, ces levures sélectionnées sont disponibles dans le commerce en grandes quantités, elles se conservent plusieurs mois, et se manipulent facilement. Si on les introduit dès le début de la fermentation, le niveau d'alcool monte rapidement, et les micro-organismes nuisibles sont éliminés.
Utilisation par l'homme
Boissons



Il y a une multiplicité indescriptible de boissons alcoolisées, dont le contenu en alcool est toujours déterminé par la fermentation alcoolique. Une condition déterminante est une matière première sucrée.
Bières
La matière première de la bière est la céréale (principalement de l'orge), riche en amidon. L'amidon n'est pas directement fermentescible, il doit être transformé en sucres pour servir de base à la fermentation, c'est le rôle du maltage. Les malteurs font germer cette céréale par trempage dans de l'eau, afin qu'elle produise certaines enzymes, puis la sèchent (touraillage). La céréale ainsi transformée s'appelle malt et peut se conserver pendant près d'un an. Les brasseurs vont ensuite empâter ce malt dans de l'eau chaude pour activer les enzymes qui dégraderont l'amidon en sucres plus simples (saccharification), notamment le maltose et permettra de les dissoudre. Après filtration et élimination des drêches, le moût ainsi obtenu sera bouilli et l'on procédera à l'ajout du houblon. Une fois refroidi, le moût sera alors ensemencé avec de la levure. Celle-ci réalisera, après consommation du dioxygène initial par respiration, une fermentation alcoolique qui décomposera les sucres issus du brassage en éthanol et dioxyde de carbone. Deux types de levures sont principalement utilisées : Saccharomyces cerevisiae pour la fermentation haute, menée autour de 20 °C, et Saccharomyces uvarum pour la fermentation basse, menée autour de 8 - 12 °C. L'éthanol reste en phase liquide, le dioxyde de carbone en partie, d'où le caractère mousseux et pétillant de la bière.
Vin
La matière première pour la fabrication du vin est le jus sucré du raisin. La levure peut fonctionner jusqu'à une concentration de 250 g/ℓ de sucre. Au-dessus, la pression osmotique est trop importante, et l'eau sort des cellules. Pour conserver l'eau autant que possible, la levure produit des solutés, principalement de la glycérine. La levure peut faire fermenter le sucre jusqu'à une concentration maximale en alcool : au-dessus, la levure meurt, parce que l'éthanol est toxique pour les cellules. La valeur limite dépend de la variété, et se situe entre 5 % et 23 %. Dès la fermentation, le contenu en éthanol protège le vin des moisissures et autres micro-organismes indésirables. Vers la fin, la fermentation est souvent maintenue en atmosphère réductrice, par isolation de l'air, afin que l'acétaldéhyde soit hydrogéné en éthanol et dioxyde de carbone. Ceci a aussi l'avantage d'empêcher l'oxydation des arômes, ainsi que l'oxydation de l'alcool en acide acétique par des bactéries. Il se dégage encore du dioxyde de carbone[16].
Les arômes issus du raisin ou arômes variétaux sont dits primaires ; ils sont dits secondaires lorsqu'issus de la fermentation ; ceux qui se développent au cours de l'élevage du vin ou de l'évolution en bouteille, sont dits tertiaires.
Mousseux
La base de la fabrication des mousseux est une deuxième fermentation alcoolique. Pour cela, on utilise un vin produit selon la méthode habituelle, on rajoute des levures et le taux de sucre est élevé soit par interruption de la première fermentation, avant complète consommation du sucre, soit par addition de sucre ou de moût[17]. Pendant cette deuxième fermentation, le taux d'alcool du vin augmente, et il se produit du dioxyde de carbone qui est maintenu dans le liquide par un bouchage hermétique[18].
Levures
Pour la production de boissons alcoolisées, on utilise des levures sélectionnées. Selon les conditions de fermentation, on choisit une souche appropriée pour obtenir le résultat souhaité. Pour protéger les matières sensibles à la chaleur, on peut conduire la fermentation à basse température (15-20 °C) par des « levures à froid. » Pour le porto et le xérès, qui atteignent un taux d'alcool jusqu'à 16 %, il faut utiliser des levures tolérantes à l'alcool. À la fin de la fermentation, le contenu final en alcool est déterminé par l'addition d'alcool neutre. Une nouveauté du marché est la « turbolevure », qui possède une tolérance à l'alcool supérieure, et peut aller dans les conditions optimales jusqu'à 20 %. Ceci ne peut être atteint en partant de jus de fruit pur. Il faut ajouter du sucre. Les turbolevures sont surtout utilisées pour faire un alcool relativement insipide, destiné à être distillé. On les utilise par exemple pour la fabrication de la vodka. On ne peut pas les utiliser pour la fabrication de liqueurs de fruits, car il est illégal d'ajouter du sucre dans la matière première pour ces liqueurs.
Spiritueux
Les boissons ayant un taux d'alcool supérieur à 20 % atteignent ce taux soit par distillation, soit par cristallisation de l'eau des matières fermentées, soit par mélange avec de l'alcool neutre (qui est lui-même obtenu par distillation ou cristallisation).
Autres aliments

Un des terrains d'application les plus importants est la boulangerie. La levure de boulanger (S. cerevisiae) est utilisée pour la fabrication de presque toutes les espèces de pains et de gâteaux traditionnels à la pâte levée, pour faire lever la pâte. Ceci se produit par la fermentation alcoolique qui dégage de fines bulles de dioxyde de carbone au sein de la pâte, et peut substantiellement en augmenter le volume. L'éthanol produit est vaporisé dans le processus de cuisson, tandis que les levures meurent à cause de la haute température.
Un aliment alcoolisé est le kéfir, fait à partir de lait. Pour le faire, on utilise des « grains de kéfir, » un mélange de levures et de bactéries vivant en symbiose. Le lactose contenu dans le lait est transformé par les bactéries en acide lactique, par fermentation lactique, puis par les levures en éthanol par fermentation alcoolique. Le kéfir a un taux d'alcool inférieur à 1 %. Chez les peuples de la steppe asiatique, on boit traditionnellement du koumis, lait de jument fermenté.
Industrie
À l'époque de la raréfaction et d'un épuisement à terme des ressources en pétrole, l'éthanol gagne de l'importance comme carburant. En outre, il est utilisé dans de nombreux procédés techniques, comme matière première pour des synthèses chimiques, ou sert à la désinfection. Cet éthanol est aussi élaboré par fermentation alcoolique par de la levure. Les matières premières sont ici des céréales à bas coût, ou des pommes de terre, dont l'amidon est décomposé en sucres par des enzymes fabriquées industriellement. Ici aussi, la fermentation ne peut pas dépasser un contenu en alcool égal à 23 %. La distillation fractionnée fournit un contenu de 96 % (azéotrope). Comme un alcool fabriqué de cette manière serait consommable, il est soumis dans la Communauté Européenne aux droits d'accise, et éventuellement à des taxes complémentaires. Une exception est faite pour l'éthanol utilisé comme carburant. De même, l'accise ne doit pas être payée pour l'alcool dénaturé, qui n'est pas consommable.
Utilisation des déchets
Le résidu solide composé par les déchets insolubles de la fabrication de l'éthanol, contient les déchets végétaux partiellement ou non hydrolysés et les levures. Il a donc une teneur accrue en protéines et peut être utilisé pour l'alimentation animale.
Notes et références
- (de) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en allemand intitulé « Alkoholische Gärung » (voir la liste des auteurs).
- (en) E. Racker, « History of the Pasteur effect and its pathobiology », Mol Cell Biochem, vol. 5, nos 1–2, , p. 17–23 (PMID 4279327, DOI 10.1007/BF01874168)
- (en) Rudolf Werner Soukup et Robert Rosner, « Scientific contributions of the first female chemists at the University of Vienna mirrored in publications in Chemical Monthly 1902–1919 », Monatshefte für Chemie - Chemical Monthly, vol. 150, no 5, , p. 961–974 (ISSN 1434-4475, DOI 10.1007/s00706-019-02408-4, lire en ligne, consulté le )
- (en) Athel Cornish-Bawden, New Beer in an Old Bottle. Eduard Buchner and the Growth of Biochemical Knowledge, Universitat de València, (ISBN 978-84-370-3328-0, lire en ligne), p. 56, 83
- (de) E. Negelein et H.J. Wulff, « Diphosphopyridinproteid, Alkohol, Acetaldehyd. », Biochemische Zeitschrift, Berlin, Springer, vol. 293, , p. 352-389 (ISSN 0366-0753)
- (en) W. Furey et al., « Structure-function relationships and flexible tetramer assembly in pyruvate decarboxylase revealed by analysis of crystal structures. », Biochimica et biophysica acta, Berlin, Springer, vol. 1385, no 2, , p. 253-270 (ISSN 0167-4889, PMID 9655915)
- (en) H. Eklund et al., « Crystallographic investigations of alcohol dehydrogenases. », EXS, Berlin, Birkhäuser, vol. 71, , p. 269-277 (ISSN 1023-294x, PMID 8032158)
- (en) P.C. Hinkle, « P/O ratios of mitochondrial oxidative phosphorylation. », Biochimica et biophysica acta, Berlin, Springer, vol. 706, nos 1-2, , p. 1-11 (ISSN 0167-4889, PMID 15620362)
- (en) J.P. van Dijken, R.A. Weusthuis et J.T. Pronk, « Kinetics of growth and sugar consumption in yeasts. », Antonie Van Leeuwenhoek. International journal of general and molecular microbiology., Dordrecht, Springer, vol. 63, nos 3-4, , p. 343-352 (ISSN 0003-6072, PMID 8279829)
- (en) K. Tonomura, « Ethanol fermentation in bacteria. », Seikagaku. The journal of Japanese Biochemical Society., Tokyo, Gakkai, vol. 59, no 10, , p. 1148-1154 (ISSN 0037-1017)
- (en) Kimmerer et McDonald, « Acetaldehyde and Ethanol Biosynthesis in Leaves of Plants. », Plant Physiology., Rockville Md, vol. 84, no 4, , p. 1204–1209 (PMCID 1056752)
- (de) Katharina Munk (dir.), Taschenlehrbuch Biologie : Mikrobiologie, Stuttgart, Thieme Verlag, , 618 p. (ISBN 978-3-13-144861-3), p. 378–379
- (en) J. Piskur et al., « How did Saccharomyces evolve to become a good brewer? », Trends in Genetics, Amsterdam, Elsevier, vol. 22, no 4, , p. 183-186 (ISSN 0168-9525, PMID 16499989)
- (en) J. Blom, M.J. De Mattos et L.A. Grivell, « Redirection of the Respiro-Fermentative Flux Distribution in Saccharomyces cerevisiae by Overexpression of the Transcription Factor Hap4p. », Applied and environmental microbiology, Washington DC, vol. 66, no 5, , p. 1970-1973 (ISSN 0099-2240, lire en ligne)
- (en) T. Conway, « The Entner-Doudoroff pathway, history, physiology and molecular biology. », Federation of European Microbiological Societies, Oxford, Blackwell, vol. 9, no 1, , p. 1-27 (ISSN 0168-6445, PMID 1389313)
- (de) Waldemar Ternes, Lebensmittel-Lexikon, Behr, , 2134 p.
- Dittrich et Großmann 2005, p. 39
- Dittrich et Großmann 2005, p. 22
- Dittrich et Großmann 2005, p. 42
Bibliographie
- (de) Gerolf Annemüller, Hans J. Manger et Peter Lietz, Die Hefe in der Brauerei. : Hefemanagement- Kulturhefe - Hefereinzucht - Hefepropagation im Bierherstellungsprozess, Berlin, Versuchs- u. Lehranstalt f. Brauerei (VLB), , 424 p. (ISBN 3-921690-50-1)
- (de) Ernst E. Bruchmann, Angewandte Biochemie. Lebensmittelchemie, Gärungschemie, Agrarchemie., Stuttgart, Ulmer, (ISBN 3-8001-2301-0) (un peu dépassé).
- (de) Helmut Hans Dittrich et Manfred Großmann, Mikrobiologie des Weines, Stuttgart, Verlag Eugen Ulmer, , 3e éd., 240 p. (ISBN 978-3-8001-4470-9)
- (de) Adam Maurizio, Geschichte der gegorenen Getränke, Sändig, (ISBN 3-253-02199-8) (Reproduction)
- (de) Lubert Stryer, Biochemie, Heidelberg, Spektrum, , 6e éd., 1224 p. (ISBN 978-3-8274-1800-5)
Voir aussi
Articles connexes
Liens externes
- (de) « Einflussfaktoren der Gärung », Wissenschaftszentrum Weihenstephan
- Collectif, Maîtrise des fermentations dirigées et spontanées, Les cahiers itinéraires d'ITV France, n° 18, , 28 p. (lire en ligne [PDF]).
 Portail de la biochimie
Portail de la biochimie  Alimentation et gastronomie
Alimentation et gastronomie  Portail de l'alcool
Portail de l'alcool  Portail de la vigne et du vin
Portail de la vigne et du vin  Portail de l’énergie
Portail de l’énergie