Viatkogorgon
Viatkogorgon ivachnenkoi

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Classe | Synapsida |
| Ordre | Therapsida |
| Sous-ordre | † Gorgonopsia |
- † Viatkogorgon ivakhnenkoi Tatarinov,
Viatkogorgon (littéralement « gorgone du Viatka ») est un genre éteint de thérapsides gorgonopsiens ayant vécu durant un stade contesté du Permien, dans ce qui est aujourd'hui la Russie européenne. Une seule espèce est connue, Viatkogorgon ivachnenkoi, décrite pour la première fois en par le paléontologue russe Leonid Petrovitch Tatarinov, et nommé en l'honneur de son collègue Mikhaïl Feodosievitch Ivakhnenko.
Le premier fossile a été découvert en dans la localité de Kotelnitch, près de la rivière Viatka, et est devenu par la suite l'holotype du taxon. Le fossile en question est d'ailleurs l'un des spécimens de gorgonopsiens les plus complets connus et comprend des éléments rarement conservés tels que des côtes abdominales ainsi qu'un anneau sclérotique (un anneau osseux situé à l'intérieur de l'œil). Un second spécimen plus grand, mais mal conservé, est également attribué à l'espèce.
Le spécimen holotype mesure environ 80 cm de long, dont 14 cm pour le crâne, faisant de Viatkogorgon un gorgonopsien relativement petit. Le second spécimen découvert est légèrement plus grand, avec un crâne long de 17 cm, et il est possible que l'holotype ait été plus jeune. L'animal possède un squelette robuste avec une position quelque peu semblable à celle d'un chien, mais avec des coudes tournés vers l'extérieur. Le museau est relevé et ses dents sont généralement recourbées vers l'arrière, pointues et dentelées. Viatkogorgon est aussi caractérisé par ses orbites anormalement grandes et son anneau sclérotique, ce qui suggère que l'animal aurait eu une mode de vie nocturne. Il se distingue des autres gorgonopsiens par le fait que l'extrémité inférieure de la barre postorbitaire est étroite et qu'il présente un très grand sillon à l'arrière de chaque côté du crâne. Le squelette de Viatkogorgon possède une colonne vertébrale avec une courbure verticale accrue, notamment dans la partie postérieure de la queue, ainsi qu'une mobilité restreinte de certains doigts des pattes arrières semblables à des nageoires, indiquant que l'animal pourrait avoir été un bon nageur. Étant donné qu'une grande majorité des gorgonopsiens ont été décrits principalement à partir de crânes, il n'est donc pas certain si les caractéristiques postcrâniennes de Viatkogorgon soient présentes chez les autres membres du groupe.
Des analyses phylogénétiques publiées en prouvent que Viatkogorgon est l'un des premiers gorgonopsiens à diverger de la base, derrière le genre Nochnitsa, également connu de Kotelnitch. Tous les autres gorgonopsiens semblent appartenir à deux groupes, russes et africains, qui divergent et apparaissent plus tard. L'âge du membre de Vanyushonki, d'où Viatkogorgon est connu, n'est pas clairement déterminé, mais il est estimé d'être daté entre la fin du Guadalupien et le début du Lopingien.
Découverte
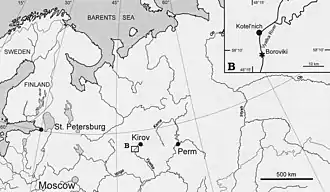
En , le paléontologue Leonid Petrovitch Tatarinov décrit un nouveau genre et une nouvelle espèce de gorgonopsiens sous le nom de Viatkogorgon ivakhnenkoi. L'holotype, sur lequel le nom scientifique est basé (catalogué PIN 2212/61 à l'Institut de Paléontologie de l'Académie des Sciences de Russie à Moscou), a été découvert en à Kotelnitch, dans l'oblast de Kirov, en Russie. La localité de Kotelnitch comprend une série de gisements datant du Permien sur les rives de la rivière Viatka[1],[2],[3].
Le nom générique Viatkogorgon fait référence à la rivière Viatka, le lieu de découverte original de l'animal, et à Gorgonops, le nom d'un genre apparenté. La qualification de « gorgone » est d'ailleurs souvent utilisé dans les noms génériques des plusieurs autres gorgonopsiens en référence aux sorcières monstrueuses de la mythologie grecque. L'épithète spécifique ivakhnenkoi honore le paléontologue russe Mikhaïl Feodosievitch Ivakhnenko, un des collègues de Tatarinov, étant à l'origine de la description de nombreux thérapsides des faunes russes[1],[3]. Ce nom spécifique est romanisé ivachnenkoi dans un article publiée en par les paléontologues russes Elena G. Kordikova et Albert J. Khlyupin, un orthographe que Tatarinov utilise également en 2004[4],[2], ce qui n'empêche pas d'autres chercheurs de continuer à utiliser l'orthographe originale[3],[5]. Les découvertes de gorgonopsiens russes commencent dans les années 1890 avec des trouvailles notables comme celui d'Inostrancevia, l'un des plus grands membres connus du groupe. Peu de découverte sont faites au cours du 20e siècle, Viatkogorgon étant le premier gorgonopsien trouvé en Russie depuis , le pays en question étant d'ailleurs le seul endroit en dehors de l'Afrique où le groupe est définitivement reconnu[1],[3].
Le squelette holotype est l'un des spécimens de gorgonopsiens les plus complets jamais découvert : il conserve la quasi-totalité du squelette postcrânien et comprend des éléments rarement conservés intacts chez les thérapsides non mammaliens, comme par exemple la présence des gastralias constituant les côtes abdominales. Le crâne est relativement mal conservé en comparaison, avec le côté gauche et le palais qui sont très endommagés. Le toit crânien est reconstruit en plâtre, à l'exception de la pointe du museau et de l'os postorbitaire gauche, un os qui forme le bord postérieur de l'orbite de l'œil. Dans l'ensemble, le crâne est également comprimé d'un côté à l'autre, ce qui le rend plus étroit qu'il n'est vu au dessus. L'anneau sclérotique droit, un anneau osseux situé à l'intérieur de l'œil, est préservé[1],[2],[3]. Ce type de détail est par ailleurs rarement conservé dans les fossiles de thériodontes, le sous-groupe de thérapsides auquel appartiennent traditionnellement les gorgonopsiens et les ancêtres des mammifères[note 2]. Le proatlas, l'atlas et l'axis, qui sont les premiers os de la colonne vertébrale, sont attachés au crâne[1],[2],[3].
Dans son article qu'il publie en , Tatarinov décrit seulement le crâne de Viatkogorgon et nomme également le nouveau genre de thérocéphale, Kotelcephalon, car l'article est limité en volume : il décrit à titre préliminaire l'arrière du crâne en [1],[2],[3]. En , Ivakhnenko signale un spécimen supplémentaire d'un Viatkogorgon de taille légèrement plus grand (catalogué PIN 4678/5), qui est très mal conservé[6]. En , les paléontologues Christian Kammerer et Vladimir Masyutin redécrivent le crâne supplémentaire de Viatkogorgon et déclarent qu'une description détaillée des restes de l'arrière du crâne améliorerait considérablement la compréhension de l'anatomie squelettique des gorgonopsiens. Ils notent cependant qu'il n'est pas disponible pour étude au cours de leurs recherches car il fait partie d'une exposition itinérante. Dans l'article, ils nomment également le nouveau gorgonopsien Nochnitsa, provenant également de Kotelnitch[3].
Description
Description simplifiée
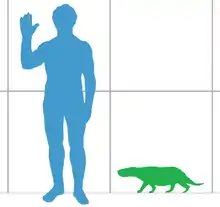
Le spécimen holotype de Viatkogorgon mesure dans les 80 centimètres de long. Le crâne mesure 14 cm, la partie conservée de la queue est d'environ 17 cm, le membre antérieur 24 cm et le membre postérieur est supérieur à 26,5 cm[2]. L'animal est relativement petit pour un gorgonopsien[3], à titre de comparaison, des genres ultérieurs tels qu’Inostrancevia ou Rubidgea mesure au moins 3 mètres de long[7]. Ivakhnenko suggère en 2002 que le spécimen holotype est peut-être un juvénile, affirmation qui est basé sur le fait qu'il possède une barre postorbitaire étroite, la largeur de cette barre s'élargissant avec l'âge chez Estemmenosuchus par exemple. Le crâne du plus grand spécimen attribué est long de 17 cm, et sa barre postorbitaire est 1 cm plus large, tandis que celui de l'holotype est de 0,6 cm. La longueur du crâne est donc multipliée par 1,3 et la largeur de la barre postorbitaire est multipliée par 1,6[6]. Les gorgonopsiens possèdent des squelettes robustes disposant des membres longs, avec une position quelque peu semblable à celle d'un chien, mais avec des coudes tournés vers l'extérieur[7]. Peu de preuves suggèrent si les thérapsides non mammaliaformes tels que les gorgonopsiens serait couverts de poils ou non[8].
Crâne
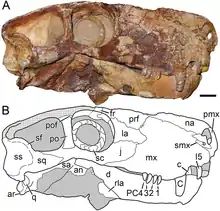
Le museau de Viatkogorgon est haut et étroit, bien qu'une grande partie de l'étroitesse du crâne soit due à la compression latérale du fossile[1]. Le prémaxillaire, l'os le plus en avant de la mâchoire supérieure, est peu exposé sur la surface latérale du museau, comme chez Nochnitsa. La barre interne qui divise les narines s'incurve légèrement vers l'arrière en vue latérale, de sorte que le bord inférieur avant du museau soit émoussé plutôt que d'être pointu. L'extrémité supérieure du prémaxillaire s'étend jusqu'au bord antérieur supérieur de la narine osseuse. Peu d'incisives supérieures sont préservées intactes dans l'holotype, mais les racines partielles et les alvéoles dentaires montrent qu'il y a cinq incisives de chaque côté, trait typique pour les gorgonopsiens. Les incisives supérieures sont faiblement courbées et spatulées, avec des dentelures claires vers leurs extrémités. Il n'est pas certain qu'elles diminuent de taille plus loin dans la rangée de dents, car les deux seules incisives intactes de l'holotype sont la même dent de chaque côté. En général, les dents sont pointues, aplaties du côté extérieur, quelque peu convexes du côté intérieur, et la zone dentelée de chaque dent est déplacée vers le bord extérieur des couronnes dentaires. Le septomaxillaire, un petit os situé entre l'os nasal et le maxillaire, a un processus latéral plus court que celle de Nochnitsa[3],[1].
Le maxillaire est proportionnellement plus haut et plus court d'avant en arrière que chez Nochnitsa, alors que le bord supérieur du maxillaire de ce dernier est arrondi, il y a un large processus qui s'étend vers l'arrière entre l'os nasal et lacrymal, l'os qui forme le bord avant de l'orbite chez Viatkogorgon. Le processus dirigé vers l'arrière qui forme le bord inférieur du maxillaire est plus court que chez Nochnitsa, n'atteignant pas le milieu de l'orbite. Le « pas » par le diastème entre les incisives et la canine est plus profond que celle de Nochnitsa, et la partie du maxillaire qui porte la canine est très convexe, ce qui donne l'impression d'une « bride » lorsqu'il est vue de côté. La canine est relativement petite pour un gorgonopsien, comme c'est le cas chez Nochnitsa, et porte des dentelures sur son bord postérieur. La rangée de dents derrière les canines sont courtes et se compose de quatre postcanines étroitement emballées, courbées vers l'arrière, contrairement à celles de Nochnitsa. Les dents postcanines sont sensiblement plus courtes que les incisives, une caractéristique régulière des gorgonopsiens, et presque trois fois plus courtes que les canines. Il y a aussi une marge faible au-dessus de la rangée de dents postcanines sur le maxillaire, dans une plus grande mesure que Nochnitsa, mais moins que chez des gorgonopsiens africains comme le genre Eriphostoma et les rubidgeinés[3],[1].
L'os nasal de Viatkogorgon est un peu plus large à l'avant par le bord postérieur de la narine osseuse que chez Nochnitsa et l'os préfrontal est proportionnellement plus court et contribue moins au bord supérieur avant de l'orbite, où l'os lacrymal forme une plus grande partie. Viatkogorgon est caractérisé par son orbite anormalement grande avec son anneau sclérotique proportionnellement grand. Le diamètre de l'orbite est de 2,8 cm, tandis que le diamètre extérieur de l'anneau sclérotique est de 2,3 cm et son diamètre intérieur 1,5 cm. L'anneau se compose de 15 osselets qui se chevauchent sans espace, et les bords de l'anneau sont régulièrement arrondis. L'os jugal, qui forme le bord inférieur de l'orbite de l'œil de Viatkogorgon, forme également une partie plus étendue du côté du visage que chez Nochnitsa[3],[1].
Viatkogorgon se distingue de tous les gorgonopsiens à l'exception de Nochnitsa en ce que l'extrémité inférieure de la barre postorbitaire, situé entre l'orbite et la fosse temporale s'ouvrant derrière cette dernière, est étroite, formant une tige droite lorsqu'elle est vue de côté. Il est plus large et n'a pas la courbure vue dans la barre de Nochnitsa. Chez d'autres gorgonopsiens, l'extrémité inférieure de la barre postorbitaire est élargie à l'endroit où elle est en contact avec l'os jugal, même si le reste de la longueur supérieure de cette barre est étroit. Le crâne de Viatkogorgon se distingue également par un très grand sillon sur l'os squamosal de chaque côté à l'arrière du crâne, qui s'étend sur une bride squamosal qui empiète sur le bord inférieur de la fenêtre temporale. En plus d'être beaucoup plus grand et expansif que le sillon squamosal de Nochnitsa, cette caractéristique a une limite distincte et une bride qui pointe vers l'arrière et vers le bas dans les deux[3].
La structure palatine de Viatkogorgon est typique des gorgonopsiens. L'animal a des dents sur l'os palatin placées sur de gros tubercules, également appelés bosses[3], avec 15 à 18 dents chacune, qui forment deux rangées qui s'étendent le long des bords externe et interne de chaque tubercule. Chaque rangée de dents forment deux lignes, la ligne extérieure étant plus longue que l'intérieure. La surface du tubercule est faiblement concave et lisse entre les dents. Il y a aussi des tubercules portant des dents sur l'os ptérygoïdien derrière l'os palatin, bien que moins développés et les dents ici (12 à 13 sur chacune) sont similaires à celles de Sauroctonus. Ce dernières sont beaucoup plus petites que les dents palatines, couvrant toute la surface des tubercules et forment trois rangées. Il y a aussi des dents sur les apophyses transverses du ptérygoïde, qui ne forment qu'une seule rangée principalement sur la partie interne, contrairement à Sauroctonus. Contrairement aux dents sur les tubercules, ces dents sont fortement usées et probablement pas remplacées chez les adultes[1].
La mâchoire inférieure de Viatkogorgon est également typique des gorgonopsiens, contrairement à celle de Nochnitsa, avec une grande symphyse mandibulaire, la zone où les deux moitiés de la mandibule se connectent à l'avant, et un menton distinct. La mâchoire inférieure est un peu plus courte que la mâchoire supérieure, indiquant un possible prognathisme. Les dents inférieures sont mal conservées dans l'holotype, mais elles semblent avoir été très similaires à la supérieure : l'incisive unique conservée est recourbée et spatulée, les dents postcanines inférieure sont faiblement recourbées et les deux ont au moins des dentelures sur leurs bords postérieurs. L'os dentaire, l'os portant la dent à l'avant de la mâchoire inférieure, est généralement plus grand que celui de Nochnitsa, et son processus coronoïde, la partie de l'endroit où la mâchoire se connecte au crâne, est plus inclinée, avec un bord postérieur faiblement concave, typique chez les gorgonopsiens. L'os articulaire est typique des gorgonopsiens et présente un processus rétro-articulaire faisant saillie vers le bas (un processus à l'arrière de la mâchoire où les muscles s'attachent)[3],[1].
Vertèbres et côtes

Étant donné que la majorités des gorgonopsiens ont été décrits principalement à partir des restes de leur crânes et avec peu de restes postcrâniens connus, les chercheurs ne savent donc pas avec certitude dans quelle mesure les si caractéristiques squelettiques de Viatkogorgon seraient répandues chez les autres membres du groupe. Viatkogorgon possède 7 vertèbres cervicales, 20 ou 21 vertèbres thoraciques du corps en avant du bassin[note 3], 3 vertèbres sacrées associées avec le bassin, et au moins 20 vertèbres caudales. La structure de l'atlas est similaire à celle des autres gorgonopsiens, et les autres vertèbres du cou sont massives, en particulier l'axis. L'axis a un intercentrum relativement grand, une très grande colonne vertébrale neurale, qui s'étend d'avant en arrière, et une face avant très haute. Les zygapophyses, les processus articulaires qui relient les vertèbres adjacentes, sont horizontales dans l'axis mais deviennent plus verticales à partir de la troisième vertèbre[2].
Les vertèbres thoraciques de Viatkogorgon sont un peu plus courtes que les vertèbres lombaires, et leurs épines neurales sont modérément hautes, avec des zygapophyses horizontales. Les épines neurales deviennent un peu plus hautes à partir du deuxième tiers de la région thoracique, et sont verticales en vue latérale, bien que dans la partie postérieure de cette région, elles sont inclinées vers l'arrière et leur bord avant deviennent convexes, montrant la transition du thoracique à lombaire vertèbres. Les 10 ou 11 côtes thoraciques antérieures sont très longues et dirigées en arrière et en bas. Ils sont étroitement accolés et attachés au bord antérieur du centrum par deux têtes articulaires rapprochées, comme dans les autres côtes. Les quatre ou cinq paires des côtes thoraciques les plus en arrière sont progressivement raccourcies, passant aux côtes lombaires, dont les têtes articulaires se rapprochent. Le sternum est long de 13 mm, en forme de plaque rectangulaire, et a trois processus robustes de chaque côté, semblable à Aelurognathus. Les gastralia de Viatkogorgon forment un cadre inhabituel en treillis de barres étroites disposées en segments, situées sous les côtes thoraciques les plus antérieures. Les barres sont disposées en deux couches à un angle l'une par rapport à l'autre, avec au moins treize barres étendues vers l'avant et vers le bas au niveau de la couche la plus profonde. Les quatre de la couche supérieure continuent les vertèbres thoraciques et s'étendent vers l'arrière et vers le bas. Les barres de gauche recouvrent les extrémités inférieures des barres de droite dans certains cas, et ces éléments sont beaucoup plus minces et plus denses que les côtes. Aucun élément reliant les côtes droite et gauche n'a été identifié[2].
La région lombaire est légèrement différenciée de la région thoracique par des côtes plus courtes. Les premières et deuxièmes côtes lombaires se courbent légèrement vers l'arrière, dirigées presque vers les côtés dans la troisième et légèrement courbées vers l'avant dans les deux vertèbres les plus en arrière. Les côtes lombaires semblent être monocéphales, contrairement aux côtes thoraciques. Les vertèbres lombaires sont massives, plus longues que les thoraciques, et avaient des zygapophyses positionnées horizontalement et des épines neurales quelque peu épaissies, dont les bords antérieurs se rétrécissent vers le haut. Les trois vertèbres lombaires les plus avant avaient des diapophyses horizontales (processus qui se projettent des côtés des arcs neuraux sur le canal neural des vertèbres), tandis que celles-ci tournent dans une direction verticale dans les deux dernières, comme on le voit chez d'autres gorgonopsiens. Des trois vertèbres sacrées, seules les deux dernières sont reliées à l'ilion du bassin par des côtes élargies, dont les extrémités se rejoignent pour former une facette articulaire commune. La facette articulaire de la vertèbre la plus antérieure est isolée des deux autres et ne sont reliée qu'à une côte rudimentaire raccourcie[2].
La région de la queue de Viatkogorgon est clairement différenciée en une partie avant et une partie arrière, la partie avant constituée de cinq vertèbres caudales qui mesurent ensemble 5,7 à 5,8 cm de longueur. Les côtes sont courtes et massives dans cette partie de la région de la queue. Les zygapophyses sot inhabituellement inclinées vers le haut, avec leurs bords postérieurs relevés par rapport à leurs bords antérieurs. La position des zygapophyses aurait restreint le mouvement latéral à la base de la queue. Les zygapophyses des quatrième et cinquième vertèbres caudales sont moins inclinées. Les côtes de deux des vertèbres caudales antérieures sont longues de 12 mm, alors qu'ils se sont raccourcis brusquement de sorte que celui de la cinquième vertèbre n'est que de 4–5 mm de long. Toutes les côtes sont à double tête et toutes les épines neurales sont relativement massives, bien qu'elles diminuent brusquement en longueur vers l'arrière. La partie postérieure de la queue est constituée de 15 vertèbres, dépourvues de côtes caudales. Dans au moins quatre de ces vertèbres, les zygapophyses sont presque surélevées à la hauteur de celles des vertèbres qui jouxtent leur devant, alors qu'elles sont positionnées presque horizontalement vers l'arrière. Les épines neurales sont basses et étroites, et l'espace entre les vertèbres a des hypapophyses bien développées (processus qui font saillie vers le bas des vertèbres) qui ne se rétrécissent pas. Les hypapophyses sont inclinées vers l'arrière et rejoignent deux vertèbres à l'avant, et couvrent les hypapophyses vers l'arrière des vertèbres adjacentes par le bas[2].
Membres et ceintures scapulaires

L'omoplate mesure de 7 cm de long, près de 1 cm large à l'articulation avec l'humérus (os du bras), et son bord supérieur élargi a une largeur de 2,5 cm. Une crête relativement basse s'étend le long du bord postérieur du tiers inférieur de l'os et légèrement déviée à l'avant du bord de l'omoplate. Les coracoïdes (partie de la ceinture scapulaire chez les vertébrés autres que les mammifères) sont déplacés quelque peu vers le haut et se rapprochent étroitement de la surface interne de l'omoplate, dépassant de l'avant et de l'arrière de cet os. Le foramen externe du coracoïde s'ouvre devant l'omoplate. Le foramen interne s'ouvre dans une incisure entre le procoracoïde et l'omoplate, une position également observée uniquement chez Gorgonops parmi les gorgonopsiens. L'humérus est plus court que le fémur (os de la cuisse), respectivement entre 9,5 et 10,3 cm de long, mais beaucoup plus massifs. La crête delto-pectorale, la où les muscles sont attachés à la partie supérieure du bras de Viatkogorgon, se projette à peu près dans la même mesure que chez Sauroctonus mais avec un grand trochanter plus peu développé. L'humérus de Viatkogorgon est similaire à celui d’Aelurognathus, bien qu'aayant une articulation du coude plus massive, presque 2 cm de diamètre à travers[2].
Les épicondyles inférieurs (les parties arrondies à l'extrémité d'un os) de l'humérus sont très bien développés et se projettent bien au-dessus des surfaces qui s'articulent avec les os inférieurs, disposant des crêtes rugueuses sur les bords. La crête de l'épicondyle médial associée au radius de l'avant-bras est massive, et la crête de l'épicondyle latéral associée à l'ulna est plus longue, 20 % de la longueur de l'humérus mais moins massive. La plupart des autres gorgonopsiens n'ont pas d'épicondyles latéraux aussi proéminents et sculptés, à l'exception d'un spécimen indéterminé. Il y a deux foramens épicondyliens, un plus étendu et en position plus haute, la différence entre les deux étant plus importante que chez Lycaenops. Les épicondyles supérieurs fortement développés le font ressembler l'humérus de Viatkogorgon en forme de S, tandis que ceux de presque tous les autres gorgonopsiens n'est que légèrement concaves d'un côté. L'avant-bras mesure environ 7 cm de long et le radius est plus étroit que le cubitus mais également aplati de haut en bas. L'ulna est beaucoup plus massif que le radius et a un processus ulnaire bien développé. Le cubitus est aplati de haut en bas et sa partie supérieure a une zone rugueuse bordée de crêtes, qui marquent l'origine des muscles fléchisseurs de la main[2].
Les pattes avant de Viatkogorgon sont étroite et mesurent environ 7 cm de long. Les doigts du milieu sont plus projetés que ceux des pattes arrière et ont une structure typique des gorgonopsiens. Viatkogorgon possède inhabituellement deux petits os centraux supplémentaires parmi les os du carpe des mains, alors que la plupart des autres gorgonopsiens n'en ont qu'un. Le cinquième doigt de la main est très spécialisé et sa structure ressemblent superficiellement à un pouce, le premier doigt présentes chez les primates (dont l'homme), les autres doigts étant d'apparence plus typique. Le premier doigt a des os courts et le troisième doigt est le plus long, comme les autres gorgonopsiens, mais sa longueur dépasse les autres dans une plus large mesure. Les os de la phalange du quatrième doigt sont complexes, leurs extrémités externes ayant un élément en forme de disque qui se fixent étroitement avec la partie principale des phalanges, mais ceux-ci sont de la même longueur que les phalanges ordinaires[2].
L'ilion de Viatkogorgon diffère de celui de Lycaenops en ce que sa partie préacétabulaire (la partie en avant de l'acétabulum, l'articulation de la hanche où le fémur se connecte) est mieux développée avec une dépression musculaire prononcée. Cela fourni probablement une zone de fixation pour le muscle iliotibial qui se connecte au bas de la jambe. Le bord supérieur de l'ilion est élevé bien au-dessus de l'endroit où cet os s'articule avec le sacrum et présente une crête longitudinale sur la surface interne. Celle-ci est séparée d'une autre crête par une fosse qui, avec une large dépression, aurait fourni une zone d'attache pour une partie du muscle ilio-pubo-fémoral interne. L'os pubien est épaissi, en forme de tige et dirigé vers l'avant et vers le bas à un angle d'environ 45 degrés, ressemblant aux os pubiens des archosauriens (le groupe qui comprend les crocodiles, les dinosaures leurs descendants oiseaux et leurs parents éteints). L'acétabulum est très grand et allongé horizontalement, mesurant plus de 20 mm de long et 15 mm élevé. Sur le bord inférieur de cette ouverture, une encoche sépare l'ischion de l'os pubien[2].
Le fémur est long de 10,3 cm, c'est-à-dire proportionnellement 70 % de la longueur du crâne. Elle est plus massive que celle de Lycaenops, et incurvée le long de son grand axe, dans la même mesure que chez Sauroctonus. La tête du fémur est ronde, environ 12 mm de diamètre, dirigé vers l'intérieur à un angle d'environ 30 degrés, et sa surface est bien construite. La tête fémorale est rétrécie vers l'arrière et elle est probablement entrée dans la partie postérieure du cotyle. Le grand trochanter descend à peu près à mi-longueur du fémur. Deux longues crêtes s'étendent le long de l'os, au-dessus et au-dessous de l'avant du grand trochanter. Le premier est probablement le trochanter interne, qui est typiquement présent chez les gorgonopsiens, mais l'autre n'est pas identifié auparavant dans le groupe. Le bas de la jambe est plus court que le fémur, 9 cm au plus, et sa sculpture de surface est plus prononcée que chez les autres gorgonopsiens, ressemblant à celles des cynodontes, qui sont les ancêtres des mammifères. Le tibia de Viatkogorgon présente des structures non identifiées chez d'autres gorgonopsiens, telles qu'une surface aplatie sur l'épicondyle préaxial et une crête dans la moitié supérieure arrière de l'os qui borde une large zone. Le péroné (os de la jambe inférieure) est plus massif que ceux des autres gorgonopsiens et a une crête sculptée massive en son milieu. Cette sculpture de surface diminue vers l'extrémité inférieure du péroné, et la crête diminue brusquement en hauteur, caractéristiques non identifiés chez d'autres gorgonopsien[2].
Le pattes arrière de Viatkogorgon mesure 7,5 cm de long et sa structure générale est similaire à celles des autres gorgonopsiens, bien que certaines caractéristiques soient notables. La rangée proximale d'os vers le talon n'est constituée que de deux os comme chez les mammifères : un talus massif et un péroné. Dans la rangée distale situé à l'avant du pied, les quatrième et cinquième tarsiens sont fusionnés, comme chez les autres gorgonopsiens, mais la rangée de centralia intermédiaire à Viatkogorgon est exceptionnellement bien développée. Les gorgonopsiens n'ont généralement qu'une seule centralia, alors que Viatkogorgon en a une supplémentaire. Une autre caractéristique inhabituelle est que les facettes articulaires des os du pied arrière s'étendent sur la face avant du péroné et du tibia de la jambe inférieure. Le tarse (le dernier groupe d'os du pied) est de 2,5 cm de long, et les métatarses (les os situés entre le tarse et les doigts) varient de 0,8 cm pour le premier à 2,1 cm pour le quatrième[2].
Les doigts de pieds sont courts, le quatrième étant de 2,5 cm, un peu plus long que le troisième. Les doigts varient légèrement en longueur, les deuxième et cinquième doigts n'étant que légèrement plus courts que les troisième et quatrième. Le raccourcissement des métatarses ajoutent à l'impression que les doigts sont courts. Les premier et cinquième doigts sont quelque peu crochus, le premier étant relativement court, mesurant environ 1,8 cm de long. La formule phalangienne des gorgonopsiens, c'est à dire le nombre et la distribution des os de la phalange dans les doigts, est similaire à celle des mammifères, tandis que l'étude de Viatkogorgon indique qu'il y a une réduction des phalanges conduisant à devenir en forme de disque combinée à leur fusion. Dans le premier doigt du pied, deux phalanges sont presque complètement fusionnées. Dans le troisième doigt, les deuxième et troisième phalanges sont raccourcies et fusionnées. Le quatrième doigt a trois phalanges raccourcies, presque en forme de disque, et fusionnées. Par conséquent, la formule phalangienne est de 1-2.3.3-4,3-5,3. Il y a un contact entre les métatarsiens vers l'arrière du pied, ainsi que des contacts entre certains métatarsiens et phalanges et des contacts entre les bases des deuxième et troisième doigts. Les ligaments médiatisent probablement les contacts antérieurs entre le métatarses et les phalanges. La phalange unguéale (os de la griffe) du cinquième doigt est remarquable du fait qu'elle est crochue et quelque peu allongée[2].
Classification


Les gorgonopsiens forment un groupe éteint de thérapsides carnivores qui comprennent les superprédateurs ayant vécu durant la fin du Permien[9],[10]. Bien qu'étants abondants, ils varient peu en morphologie. Répandus principalement en Afrique australe durant cette période, ils sont peu représentés dans les archives fossiles du reste du monde et ne sont définitivement connus que de Russie en dehors de l'Afrique[3]. Les gorgonopsiens incarnent les premiers prédateurs à développer des « dents de sabre », bien avant que les premiers mammifères et dinosaures n'apparaissent[10]. Cette caractéristique a ensuite évolué indépendamment et plusieurs fois dans différents groupes de mammifères prédateurs, tels que les félins machairodontes ou les thylacosmilidés[11]. Ils s'éteignent lors de l'extinction Permien-Trias[7],[10].
Dans sa description publié en , Tatarinov classe Viatkogorgon comme appartenant à la sous-famille de gorgonopsiens Sycosaurinae au sein de la famille des Gorgonopsidae, sur la base de caractéristiques telles que la petite taille et l'étroitesse du museau. Cette sous-famille n'est alors pas connue auparavant en Russie. Il note également quelques similitudes avec les membres de la famille des Rubidgeidae[1]. Ivakhnenko considère Sycosaurinae, y compris Viatkogorgon, comme faisant partie des Rubidgeidae en [6]. Dans sa description de du squelette postcrânien, Tatarinov averti que cela fournit peu d'informations sur la position taxonomique de Viatkogorgon parmi les gorgonopsiens, certaines caractéristiques étant uniques et d'autres apparaissant chez d'autres membres du groupe. Le manque d'information résultant du fait que peu de gorgonopsiens ont été découverts avec des squelettes complets. Néanmoins, il trouve la structure des pattes de Viatkogorgon similaire à celle des gorgonopsiens africains tels qu'Arctognathus et Aelurognathus[1].
En , Kammerer et Masyutin déclare qu'en raison de l'incertitude concernant l'évolution précoce des gorgonopsiens, Viatkogorgon et Nochnitsa élargissent donc la connaissance de ce groupe du Permien moyen et/ou du Permien supérieur de la Laurasie, la masse continentale nord du supercontinent Pangée, composé de ce qui est maintenant l'Eurasie et l'Amérique du Nord. Selon les analyses phylogénétiques, Nochnitsa s'avère être le gorgonopsien le plus basal suivi de Viatkogorgon, en raison de son absence d'expansion inférieure sur la barre postorbitaire, une caractéristique observée dans les genres divergents plus tard, ceux-ci étant en dehors d'un clade regroupant tous les autres gorgonopsiens. Ce groupe est par la suite de nouveau divisé en deux clades, l'un composé des gorgonopsiens russes et l'autre des gorgonopsiens africains, sur la base de caractéristiques partagées sur les crânes[3],[10].
Le cladogramme suivant montre la position phylogénétique de Viatkogorgon au sein du taxon Gorgonopsia selon une étude publié par Eva-Maria Bendel et al. en [10], qui suit en grande partie le cladogramme établi par Kammerer et Masyutin quelque mois plutôt[3] :
| ◄ Gorgonopsia |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
En revanche, les analyses précédentes ne trouve pas que les gorgonopsiens sont à regrouper géographiquement, certaines études plaçant des genres russes, dont Inostrancevia, près ou dans des taxons africains, personne soupçonnant que différents groupes de gorgonopsiens seraient endémiques dans différentes régions. Kammerer et Masyutin trouve cela particulier, car il existe de nombreux taxons frères russo-africains dans d'autres groupes de thérapsides, notamment les dicynodontes et les burnétiamorphes. Cela indique qu'il y aurait eu une vaste dispersion de groupes de thérapsides contemporains entre les continents. Ils avertissent que la paléobiogéographie des tétrapodes pendant le Permien reste mal comprise, les capacités de dispersion attendues de divers groupes de thérapsides différent souvent de ce qui peut être vu dans les archives fossiles et suggèrent que des recherches supplémentaires sont nécessaires[3].
Kammerer et Masyutin note que si Viatkogorgon et Nochnitsa ajoutent à la diversité des gorgonopsiens de la faune de Kotelnitch, le groupe reste moins riche en espèces que les thérocéphales de cette région. La faible diversité et la petite taille des gorgonopsiens indique que l'assemblage de thérapsides est similaire à celui observé dans la zone d'assemblage de Pristerognathus (en), dans le bassin du Karoo, en Afrique du Sud, avant le cycle principal de diversification des gorgonopsiens. À cet égard, ils trouvent la position basale de Nochnitsa et Viatkogorgon intrigante, bien qu'ils ne trouvent pas indicative de la localité de Kotelnitch, étant d'un âge plus précoce que les strates sud-africaines du Permien moyen qui contient des fossiles de gorgonopsiens, sur la base d'autres types de groupes de thérapsides trouvés parmi ces faunes. Ils déclarent que si le clade russe des gorgonopsiens aurait probablement divergé au moment où la faune de Kotelnitch existait, l'absence de ce clade dans cette localité suggère qu'ils n'ont pas encore subi de diversification substantielle en Russie, et ne sont devenus que le groupe dominant de thérapsides prédateurs dans la région plus tard[3].
Paléobiologie
Mode de vie

En , Tatarinov note que Viatkogorgon aurait pu presser la nourriture contre les zones palatines concaves et lisses entre les dents palatines avec la langue[1]. Les dents palatines aurait aidé les premiers synapsides et autres amniotes basaux à saisir la nourriture : ceux-ci sont absents chez les mammifères qui ont des palais secondaires à la place[12]. En , Kammerer et Masyutin considèrent que l'anneau sclérotique de Viatkogorgon est remarquablement grand par rapport à son orbite, tombant bien dans ce qui serait considéré comme une vision scotopique, ce qui suggère des habitudes nocturnes[3]. Le paléontologue russe Valeriy Golubev examine des assemblages de vertébrés terrestres datant du Permien dans l'Europe de l'Est en : il note que Viatkogorgon et le thérocéphale Viatkosuchus, les plus grands prédateurs du sous-assemblage de Kotelnitch, sont relativement petits et pas très différents en taille des prédateurs plus petits de leur assemblage, tels que les thérocéphales Scalopodon (en), Scalopodontes et Kotelcephalon[13]. En , les paléontologues russes Yulia A. Suchkova et Golubev déclare que le thérocéphale Gorynychus, découvert également à Kotelnitch, aurait partagé sa niche de prédateur dominant avec Viatkogorgon[5].
Le paléoartiste et écrivain espagnol Mauricio Antón donne un aperçu de la biologie des gorgonopsiens dans un livre qu'il publie en , écrivant qu'en dépit de leurs différences avec les mammifères dit à « dents de sabre », de nombreuses caractéristiques de leurs squelettes indiquent qu'ils auraient été des prédateurs actifs. Alors que leurs cerveaux sont proportionnellement plus petit que celui des mammifères et que leurs yeux placés sur le côté offrent une vision stéréoscopique limitée, ils possèdent des cornets bien développés dans leur cavité nasale, une caractéristique associée à un odorat avancé qui les aurait aidés pour traquer leurs proies. Les canines auraient été utilisées pour délivrer une morsure tranchante, tandis que les incisives, formant un arc devant les canines, retiendrait la proie en coupant la chair tout en se nourrissant. Pour leur permettre d'augmenter leur ouverture lorsqu'ils mordent, les gorgonopsiens possèdent plusieurs os dans leurs mandibules qui peuvent se déplacer les uns par rapport aux autres et qui auraient une double articulation avec le crâne, contrairement aux mammifères où l'os articulaire de l'articulation arrière constitue l'un de os de l'oreille : le marteau[14]. Antón imagine que les gorgonopsiens chasseraient en quittant leur abri lorsque la proie serait suffisamment proche d'eux. Il utiliseraient leur vitesse relativement plus grande pour bondir rapidement dessus, les attraper avec leurs membres antérieurs et mordre toute partie du corps qui tiendrait dans leurs mâchoires. Une telle morsure entraînerait une perte de sang importante et le prédateur continuerait à essayer de mordre les parties vulnérables du corps[15].
Déplacement
En , Tatarinov interprète le comportement de Viatkogorgon en fonction de ses caractéristiques squelettiques, qui, bien qu'étant généralement similaires à celles des autres gorgonopsiens, présentent certaines caractéristiques uniques ou mal comprises. Celles-ci concernent en particulier des attributs de l'appareil locomoteur, qui indiquent un stade d'adaptations à la nage, tandis que d'autres caractéristiques qui sont cohérentes avec celles couramment observées dans son groupe. Comme les autres gorgonopsiens, l'animal possède une longue région lombaire des vertèbres, dont les vertèbres les plus postérieures présentent une capacité accrue de courbure verticale, ce qui est également le cas pour les vertèbres du cou et, exceptionnellement, la partie postérieure de la région de la queue[2].
La présence de gastralia bien développées est l'une des caractéristiques les plus inhabituelles chez les thériodontes, tout comme les hypophyses de la région de la queue. De plus, le pied large est quelque peu raccourci avec des contacts intermétatarsiens inhabituels, qui limitent la mobilité des doigts individuels, indiquant la transformation des pattes en une structure semblable à une nageoire. Tatarinov émet l'hypothèse que ces caractéristiques seraient des adaptations pour la natation : alors que Viatkogorgon n'est pas un prédateur aquatique spécialisé, Tatarinov suggère que sa queue et ses pattes lui permettraient de mieux nager que la plupart des autres gorgonopsiens. Il pense en particulier que la partie postérieure de la queue serait probablement utilisée pour nager, comme c'est le cas des reptiles à longue queue qui ont des hypapophyses, comme les célèbres mosasaures. Tatarinov note que puisque la griffe du cinquième doigt du membre postérieur est crochue et quelque peu allongée, ce dernier aurait pu être utilisée pour la protection et le toilettage, comme c'est le cas chez les monotrèmes actuels. Le premier doigt aurait pu avoir cette fonction à un degré moindre, car il n'est pas en contact avec les os des pattes arrières. Les membres antérieurs de Viatkogorgon sont moins spécialisé et auraient pu remplir des fonctions plus importantes, car il manque de contacts entre les doigts[2].
Antón déclare en que même si les squelettes postcrâniens des gorgonopsiens sont fondamentalement de forme reptiliennes, leurs positions seraient beaucoup plus droite que chez les synapsides plus primitifs, comme les pélycosaures, qui sont plus tentaculaires. La locomotion régulière des gorgonopsiens serait similaire aux « marches hautes » observées chez les crocodiliens, dans lesquelles le ventre est porté au-dessus du sol, les pieds pointés vers l'avant et les membres portés sous le tronc plutôt que sur les côtés. Les membres antérieurs ont une posture plus horizontale que les membres postérieurs, avec les coudes pointés vers l'extérieur pendant le mouvement, mais la démarche des membres postérieurs aurait ressemblé à celle des mammifères. Comme chez les reptiles, les muscles de la queue sont importants dans la flexion du membre postérieur, alors que les queues des mammifères sont simplement utilisé pour l'équilibre. Leurs pattes seraient probablement plantigrades, c'est-à-dire que les semelles sont placées à plat sur le sol, et seraient probablement plus rapides et agiles que celui leurs proies. Leurs pieds sont plus symétriques par rapport à la condition reptilienne, rendant le contact avec le sol plus efficace, semblable à celui de certains mammifères quand ils courent[15].
Paléoenvironnement
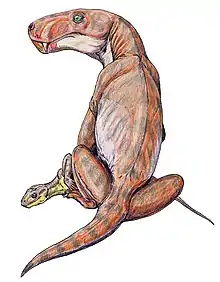
Viatkogorgon est connu de la localité de Kotelnitch, qui consiste en une série d'expositions de lits rouges datant du Permien le long des rives de la rivière Viatka, situé dans le nord de la Russie. Il provient spécifiquement du membre de Vanyushonki, qui est la plus ancienne unité rocheuse de la succession de Kotelnitch, composé de mudstones pâles ou bruns (composée d'argile et de limons, avec un peu de sable à grain fin) et de mudstones gris ou rouge foncé à la base de cette exposition. Ces mudstones ont peut-être été déposés en suspension dans des plans d'eau stagnants sur des plaines inondables ou des lacs éphémères peu profonds, qui restent inondés pendant de courtes périodes, mais l'environnement exact n'est pas encore déterminé en raison de l'absence de structures primaires des sédiments. L'abondance de radicelles fossiles et de grands herbivores indique que le paysage représenté par le membre de Vanyushonki serait relativement humide et bien végétalisé. Le complexe faunique de Kotelnitch est peut-être du même âge à celles retrouvés en Afrique du Sud, qui date soit de la dernière période du Permien moyen (c'est-à-dire 260,26 millions d'années avant notre ère), soit du début du Permien supérieur[3],[16].
Le membre de Vanyushonki est la source de la plupart des fossiles de tétrapodes de la localité de Kotelnitch, avec des restes squelettiques abondants, souvent constitués de squelettes articulés complets. En dehors des gorgonopsiens Viatkogorgon et Nochnitsa, les thérapsides de la localité sont les anomodontes Suminia et plusieurs genres de thérocephales, dont Chlynovia (en), Gorynychus, Karenites (en), Perplexisaurus, Scalopodon (en), Scalopodontes et Viatkosuchus. Des parareptiles comme Deltavjatia et Emeroleter y sont particulièrement abondant[3],[17]. Des ostracodes fossiles, des traces de racines et des souches d'arbres sont également trouvables[16].
Notes et références
Notes
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Viatkogorgon » (voir la liste des auteurs).
- Ce taxon a été romanisée en sur la base de l'épithète spécifique ivakhnenkoi avant d'être plus tard utilisée par l'auteur original de la description de l'animal en et par d'autre chercheurs ultérieurement, pour plus d'information, voir section « Découverte ».
- La validité du taxon Theriodontia est débattu. Bien qu'un nombre important d'études reconnaissent l'existence d'un possible clade réunissant le dernier ancêtre commun des gorgonopsiens et des euthériodontes (le groupe qui inclut les mammifères, leurs ancêtres et une lignée éteinte apparenté), des recherches supplémentaires plus récentes se montrent septique quand à cette proposition[3].
- 15 en excluant les cinq vertèbres lombaires, celles entre la cage thoracique et le bassin[2].
Références
- (en) Leonid P. Tatarinov, « New theriodonts (Reptilia) from the Late Permian fauna of the Kotelnich locality, Kirov Region », Paleontological Journal, vol. 5, no 33, , p. 550-554.
- (en) Leonid P. Tatarinov, « A postcranial skeleton of the gorgonopian Viatkogorgon ivachnenkoi (Reptilia, Theriodontia) from the Upper Permian Kotelnich locality, Kirov Region », Paleontological Journal, vol. 38, no 4, , p. 437-447.
- (en) Christian F. Kammerer et Vladimir Masyutin, « Gorgonopsian therapsids (Nochnitsa gen. nov. and Viatkogorgon) from the Permian Kotelnich locality of Russia », PeerJ, vol. 6, , e4954 (PMID 29900078, PMCID 5995105, DOI 10.7717/peerj.4954
 ).
). - (en) Elena G. Kordikova et J. Khlyupin Albert, « First evidence of a neonate dentition in pareiasaurs from the Upper Permian of Russia », Acta Palaeontologica Polonica, vol. 46, no 4, , p. 589-594 (lire en ligne).
- (en) Yulia A. Suchkova et Valeriy K. Golubev, « A new Permian therocephalian (Therocephalia, Theromorpha) from the Sundyr assemblage of Eastern Europe », Paleontological Journal, vol. 53, no 4, , p. 411–417 (DOI 10.1134/S0031030119040117, S2CID 201659515, lire en ligne)
- (en) Mikhail F. Ivakhnenko, « Taxonomy of East European Gorgonopia (Therapsida) », Paleontological Journal, vol. 36, no 3, , p. 283-292 (ISSN 0031-0301).
- Antón 2013, p. 79-81.
- (en) Julien Benoit, Paul R. Manger et Bruce S. Rubidge, « Palaeoneurological clues to the evolution of defining mammalian soft tissue traits », Scientific Reports, vol. 6, no 1, , p. 25604 (PMID 27157809, PMCID 4860582, DOI 10.1038/srep25604 , Bibcode 2016NatSR...625604B).
- Antón 2013, p. 7-23.
- (en) Eva-Maria Bendel, Christian F. Kammerer, Nikolay Kardjilov, Vincent Fernandez et Jörg Fröbisch, « Cranial anatomy of the gorgonopsian Cynariops robustus based on CT-reconstruction », PLOS ONE, vol. 13, no 11, , e0207367 (PMID 30485338, PMCID 6261584, DOI 10.1371/journal.pone.0207367 , Bibcode 2018PLoSO..1307367B)
- Antón 2013, p. 7-22.
- (en) Ryoko Matsumoto et Susan E. Evans, « The palatal dentition of tetrapods and its functional significance », Journal of Anatomy, vol. 230, no 1, , p. 47–65 (PMID 27542892, PMCID 5192890, DOI 10.1111/joa.12534 ).
- (en) Valeriy K. Golubev, « The faunal assemblages of Permian terrestrial vertebrates from Eastern Europe », Paleontological Journal, vol. 34, no 2, , p. 211-224 (lire en ligne).
- (en) Aitor Navarro‐Díaz, Borja Esteve‐Altava et Diego Rasskin‐Gutman, « Disconnecting bones within the jaw‐otic network modules underlies mammalian middle ear evolution », Journal of Anatomy, vol. 235, no 1, , p. 15–33 (PMID 30977522, PMCID 6579944, DOI 10.1111/joa.12992 ).
- Antón 2013, p. 204-209.
- (en) Michael J. Benton, Andrew J. Newell, Al'bert Y. Khlyupin, Il'ya S. Shumov, Gregory D. Price et Andrey A. Kurkin, « Preservation of exceptional vertebrate assemblages in Middle Permian fluviolacustrine mudstones of Kotel'nich, Russia: stratigraphy, sedimentology, and taphonomy », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 319-320, , p. 58–83 (DOI 10.1016/j.palaeo.2012.01.005, Bibcode 2012PPP...319...58B, S2CID 129753042, lire en ligne [PDF])
- (en) Christian F. Kammerer et Vladimir Masyutin, « A new therocephalian (Gorynychus masyutinae gen. et sp. nov.) from the Permian Kotelnich locality, Kirov Region, Russia », PeerJ, vol. 6, , e4933 (PMID 29900076, PMCID 5995100, DOI 10.7717/peerj.4933 ).
Voir aussi
Articles connexes
Bibliographie
![]() : document utilisé comme source pour la rédaction de cet article.
: document utilisé comme source pour la rédaction de cet article.
- (en) Mauricio Antón, Sabertooth, Bloomington, Indiana University Press, (ISBN 978-0-253-01042-1, OCLC 857070029, lire en ligne).

Liens externes
- Ressources relatives au vivant :
 Portail de la paléontologie
Portail de la paléontologie  Portail de la Russie
Portail de la Russie