Alpha-neurotoxin
α-Neurotoxins are a group of neurotoxic peptides found in the venom of snakes in the families Elapidae and Hydrophiidae. They can cause paralysis, respiratory failure, and death. Members of the three-finger toxin protein family, they are antagonists of post-synaptic nicotinic acetylcholine receptors (nAChRs) in the neuromuscular synapse that bind competitively and irreversibly, preventing synaptic acetylcholine (ACh) from opening the ion channel. Over 100 α-neurotoxins have been identified and sequenced.[2]
History
The term α-neurotoxin was coined by C.C. Chang, who designated the postsynaptic bungarotoxin with the α- prefix because it happened to be slowest moving of the bungarotoxins under starch zone electrophoresis.[3] The "α-" prefix subsequently came to connote any toxins with postsynaptic action. Members of this group are sometimes referred to as "curaremimetics" due to the similarity of their effects with the plant alkaloid curare.[4][5]
As more snake venoms were characterized, many were found to contain homologous nAChR-antagonist proteins. These came to be collectively known as the snake venom α-neurotoxins.[5]
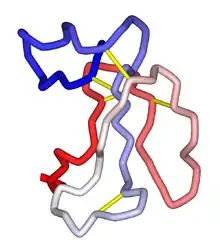
General structure
All α-neurotoxins share the three-finger toxin tertiary structure, consisting of a small globular core containing four disulfide bonds, three loops or "fingers", and a C-terminal tail.[4] The class can be divided into two groups distinguished by length; short-chain neurotoxins have 60-62 residues and only the four core disulfide bonds characteristic of the fold, while long-chain neurotoxins have 66 or more residues, often including a longer C-terminus, and an additional disulfide bond in the second "finger" loop.[4][6] These classes have significant sequence homology and share the same three-dimensional structure, but have differing specificities and kinetics of association/dissociation with the receptor.[7] Localized mobility at the tips of fingers I and II is essential for binding.[8] Accordingly, mutation of these residues produces large effects on binding.[9][6] The additional disulfide bond in the second loop of the long-chain forms is likewise thought to influence binding specificity.[4] Although both short and long-chain neurotoxins bind the same site on their target receptors, short-chain neurotoxins do not potently block α7 homo-oligomeric neuronal AChRs,[10] while long-chain neurotoxins do.[4] α-bungarotoxin and α-cobratoxin are both long-type.[6]
Functions
For specifics, see Alpha-Bungarotoxin and nicotinic acetylcholine receptor
α-Neurotoxins antagonistically bind tightly and noncovalently to nAChRs of skeletal muscles, thereby blocking the action of ACh at the postsynaptic membrane, inhibiting ion flow and leading to paralysis. nAChRs contain two binding sites for snake venom neurotoxins. Some computational studies of the mechanism of inhibition using normal mode dynamics[11] suggest that a twist-like motion caused by ACh binding may be responsible for pore opening, and that this motion is inhibited by toxin binding.[11][12]
Evolution
Although three-finger protein domains are widespread, three-finger toxins appear only in snakes, and are particularly enriched in elapids.[13] There is evidence that alpha-neurotoxins have evolved rapidly and are subject to positive selection,[14] possibly due to an evolutionary arms race with prey species.[15]
Snake nAchRs have specific sequence features that render them poor binding partners for alpha-neurotoxins.[16][17] Some mammalian lineages also display mutations conferring resistance to alpha-neurotoxins; such resistance is believed to have evolved convergently at least four times in mammals, reflecting two different biochemical mechanisms of adaptation.[18] The introduction of glycosylation sites on the receptor, resulting in steric hindrance at the neurotoxin binding site, is a well-characterized resistance mechanism found in mongooses, while the honey badger, domestic pig, and hedgehog lineages replace aromatic amino acids with charged residues; at least in some lineages, these molecular adaptations likely reflect predation on venomous snakes.[18][16]
References
- Zeng H, Moise L, Grant MA, Hawrot E (June 2001). "The solution structure of the complex formed between alpha-bungarotoxin and an 18-mer cognate peptide derived from the alpha 1 subunit of the nicotinic acetylcholine receptor from Torpedo californica". The Journal of Biological Chemistry. 276 (25): 22930–40. doi:10.1074/jbc.M102300200. PMID 11312275.
- Hodgson WC, Wickramaratna JC (September 2002). "In vitro neuromuscular activity of snake venoms". Clinical and Experimental Pharmacology & Physiology. 29 (9): 807–14. doi:10.1046/j.1440-1681.2002.03740.x. PMID 12165047. S2CID 20158638.
- Chang CC (1999). "Looking back on the discovery of alpha-bungarotoxin". Journal of Biomedical Science. 6 (6): 368–75. doi:10.1159/000025412. PMID 10545772. S2CID 84443027.
- Kini RM, Doley R (November 2010). "Structure, function and evolution of three-finger toxins: mini proteins with multiple targets". Toxicon. 56 (6): 855–67. doi:10.1016/j.toxicon.2010.07.010. PMID 20670641.
- Barber CM, Isbister GK, Hodgson WC (May 2013). "Alpha neurotoxins". Toxicon. 66: 47–58. doi:10.1016/j.toxicon.2013.01.019. PMID 23416229.
- Moise L, Piserchio A, Basus VJ, Hawrot E (April 2002). "NMR structural analysis of alpha-bungarotoxin and its complex with the principal alpha-neurotoxin-binding sequence on the alpha 7 subunit of a neuronal nicotinic acetylcholine receptor". The Journal of Biological Chemistry. 277 (14): 12406–17. doi:10.1074/jbc.M110320200. PMID 11790782.
- Tsetlin V (September 1999). "Snake venom alpha-neurotoxins and other 'three-finger' proteins". European Journal of Biochemistry. 264 (2): 281–6. doi:10.1046/j.1432-1327.1999.00623.x. PMID 10491072.
- Connolly PJ, Stern AS, Hoch JC (January 1996). "Solution structure of LSIII, a long neurotoxin from the venom of Laticauda semifasciata". Biochemistry. 35 (2): 418–26. doi:10.1021/bi9520287. PMID 8555211.
- Trémeau O, Lemaire C, Drevet P, Pinkasfeld S, Ducancel F, Boulain JC, Ménez A (April 1995). "Genetic engineering of snake toxins. The functional site of Erabutoxin a, as delineated by site-directed mutagenesis, includes variant residues". The Journal of Biological Chemistry. 270 (16): 9362–9. doi:10.1074/jbc.270.16.9362. PMID 7721859.
- de la Rosa G, Corrales-García LL, Rodriguez-Ruiz X, López-Vera E, Corzo G (July 2018). "Short-chain consensus alpha-neurotoxin: a synthetic 60-mer peptide with generic traits and enhanced immunogenic properties". Amino Acids. 50 (7): 885–895. doi:10.1007/s00726-018-2556-0. PMID 29626299. S2CID 4638613.
- Levitt M, Sander C, Stern PS (February 1985). "Protein normal-mode dynamics: trypsin inhibitor, crambin, ribonuclease and lysozyme". Journal of Molecular Biology. 181 (3): 423–47. doi:10.1016/0022-2836(85)90230-X. PMID 2580101.
- Samson AO, Levitt M (April 2008). "Inhibition mechanism of the acetylcholine receptor by alpha-neurotoxins as revealed by normal-mode dynamics". Biochemistry. 47 (13): 4065–70. doi:10.1021/bi702272j. PMC 2750825. PMID 18327915.
- Kessler P, Marchot P, Silva M, Servent D (August 2017). "The three-finger toxin fold: a multifunctional structural scaffold able to modulate cholinergic functions". Journal of Neurochemistry. 142 Suppl 2: 7–18. doi:10.1111/jnc.13975. PMID 28326549.
- Sunagar, Kartik; Jackson, Timothy N. W.; Undheim, Eivind A. B.; Ali, Syed A.; Antunes, Agostinho; Fry, Bryan G. (November 2013). "Three-Fingered RAVERs: Rapid Accumulation of Variations in Exposed Residues of Snake Venom Toxins". Toxins. 5 (11): 2172–2208. doi:10.3390/toxins5112172. PMC 3847720. PMID 24253238.
- Casewell NR, Wüster W, Vonk FJ, Harrison RA, Fry BG (April 2013). "Complex cocktails: the evolutionary novelty of venoms". Trends in Ecology & Evolution. 28 (4): 219–29. doi:10.1016/j.tree.2012.10.020. PMID 23219381.
- Arbuckle K, Rodríguez de la Vega RC, Casewell NR (December 2017). "Coevolution takes the sting out of it: Evolutionary biology and mechanisms of toxin resistance in animals" (PDF). Toxicon. 140: 118–131. doi:10.1016/j.toxicon.2017.10.026. PMID 29111116. S2CID 11196041.
- Neumann D, Barchan D, Horowitz M, Kochva E, Fuchs S (September 1989). "Snake acetylcholine receptor: cloning of the domain containing the four extracellular cysteines of the alpha subunit". Proceedings of the National Academy of Sciences of the United States of America. 86 (18): 7255–9. Bibcode:1989PNAS...86.7255N. doi:10.1073/pnas.86.18.7255. PMC 298036. PMID 2780569.
- Drabeck DH, Dean AM, Jansa SA (June 2015). "Why the honey badger don't care: Convergent evolution of venom-targeted nicotinic acetylcholine receptors in mammals that survive venomous snake bites". Toxicon. 99: 68–72. doi:10.1016/j.toxicon.2015.03.007. PMID 25796346.