Implantation (embryology)
Implantation (also known as nidation) is the stage in the embryonic development of mammals in which the blastocyst hatches as the embryo, adheres, and invades into the wall of the female's uterus.[1] Implantation is the first stage of gestation, and when successful the female is considered to be pregnant. The implanted embryo will receive oxygen and nutrients in order to grow.
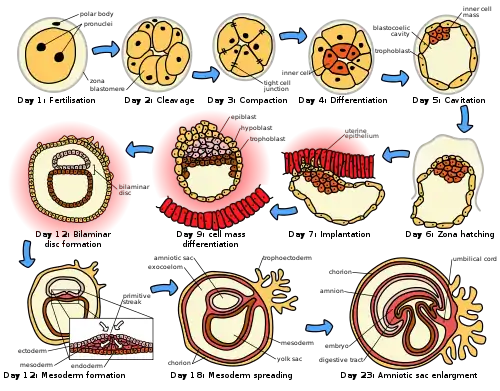
There is an extensive variation in the type of trophoblast cells, and structures of the placenta across the different species of mammals.[2] Of the five recognised stages of implantation including two pre-implantation stages that precede placentation, the first four are similar across the species. The five stages are migration and hatching, pre-contact, attachment, adhesion, and invasion.[2] The two pre-implantation stages are associated with the pre-implantation embryo.[3][4]
In humans following the stage of hatching that takes place around four to five days after fertilization, the process of implantation begins. By the end of the first week the blastocyst is superficially attached to the uterine endometrium. By the end of the second week implantation has completed.[5]
Implantation stages
There are five recognized stages of implantation in mammals, including two pre-implantation stages that precede the formation of the placenta. They are: migration and hatching, pre-contact, attachment, adhesion, and invasion. The first four stages are similar across the species with the process of invasion being variable.[2][6] These three stages of apposition, attachment, and invasion are also alternatively termed contact (apposition), adhesion (attachment), and penetration (invasion),[4][3] and can only take place during a limited timeframe known as the window of implantation.
Migration and hatching
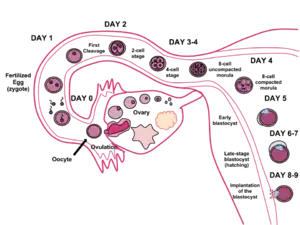
There are two stages of migration involved in implantation, the first is the migration of the zygote, and the second is the migration of the trophoblast.[7]
Following the fertilization of the oocyte that creates the single diploid cell called the zygote, the zygote starts to migrate towards the uterus. Fertilization takes place in the ampulla of the fallopian tube, and the zygote is transported along the tube to the uterus. The fallopian tube is lined with ciliated epithelium, and it is the cilia that move the zygote.[7]
During this migration the zygote undergoes a number of cell divisions that creates a ball of 16 compacted blastomeres called a morula.[8] The morula enters the uterus after three or four days, and as it does a cavity called the blastocoel is formed in the morula to produce the blastocyst. The blastocyst contains the inner cell mass that will go on to develop into the embryo proper, and an outer cell layer of trophoblasts that will develop into the extraembryonic membranes (fetal membranes).[9]
The blastocyst is still enclosed in the egg-coat known as the zona pellucida, and for it to be able to implant into the uterine wall it must rid itself of this covering. This stage is known as zona hatching, and when there is sufficient dissolution the blastocyst is able to initiate the apposition stage of implantation. Lytic factors in the uterine cavity, as well as factors from the blastocyst itself are essential for the breakdown of the egg-coat. Mechanisms in the latter are indicated by the fact that the zona pellucida remains intact if an unfertilized egg is placed in the uterus under the same conditions.[10]
Among the known molecular regulators that promote hatching are predominantly proteases that are stimulated by various growth factors.[11] The blastocyst also produces cytokines, both pro-inflammatory and anti-inflammatory, that have crucial roles during implantation and other stages of pregnancy. Both types of cytokines modulate the activity of proteases, including MMPs, plasminogen activators, and cathepsins.[11] It is unknown whether the cytokines involved in hatching are pro-inflammatory or anti-inflammatory, or which proteases are involved. However, it is well accepted that the pro-inflammatory cytokines are dominant during implantation. Cytokines are also present in the uterine milk which might regulate the development and function of the blastocyst but there is no evidence to support their involvement in hatching. Leukemia inhibitory factor (LIF) is a pro-inflammatory cytokine expressed in the endometrium during the luteal phase of the menstrual cycle, with the highest expression seen during the window of implantation. LIF plays a role in adhesion and invasion. [11]
Assisted zona hatching may take place in assisted reproduction, where the zona pellucida may be artificially pierced to facilitate hatching.[12]
Apposition
Following zona hatching, the very first loose connection or contact between the blastocyst and the endometrium is called the apposition. The apposition is usually made where there is a small crypt in the endometrium, and also where there has been enough breakdown of the zona pellucida to allow the blastocyst trophoblast to directly contact the underlying endometrium. Ultimately, the inner cell mass (also embryoblast), inside the trophoblast layer, is aligned closest to the decidua. If the inner cell mass is not aligned with the decidua at apposition, it has the ability to freely rotate within the trophoblast and achieve this alignment.[13]
Molecular mechanism
The identity of the molecules on the trophoblast and the endometrial epithelia that mediate the initial interaction between the two remain unidentified. However, a number of research groups have proposed that MUC1, a member of the mucin family of glycosylated proteins, is involved.[14] MUC1 is a transmembrane glycoprotein expressed at the apical surface of endometrial epithelial cells during the window of implantation in humans and has been shown to be differentially expressed between fertile and infertile subjects during this time.[14] MUC1 displays carbohydrate moieties on its extracellular domain that are ligands of L-selectin, a cell adhesion molecule on the surface of trophoblast cells.[15][16] An in vitro model of implantation gave evidence to support the hypothesis that L-selectin mediates apposition of the blastocyst to the uterine epithelium by interacting with its ligands.[17]
Adhesion
Adhesion is a much stronger attachment to the endometrium than the loose apposition.
The trophoblasts adhere by penetrating the endometrium, with protrusions of trophoblast cells.
This adhering activity is by microvilli that are on the trophoblast. The trophoblast have binding fiber connections, laminin, collagen type IV, and integrins that assist in this adhesion process.[18]
Mucin-16 is a transmembrane mucin expressed at the apical surface of uterine epithelia. This mucin prevents the blastocyst from implanting in an undesired located on the epithelium. Thus, MUC-16 inhibits cell-cell adhesion. Its removal during pinopode formation has been shown to facilitate trophoblast invasion in vitro.[19]
Invasion
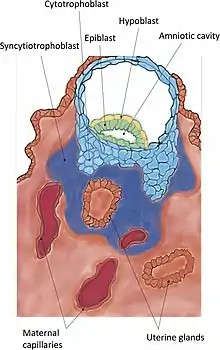
Invasion is the further establishment of the blastocyst into the endometrium. The protrusions of trophoblast cells that adhere into the endometrium continue to proliferate and penetrate into the endometrium. As these trophoblasts penetrate, they terminally differentiate into a multinucleated tissue, a syncytium known as the syncytiotrophoblast. Between this layer and the blastocyst lies the cytotrophoblast.[20] When the syncytiotrophoblast reaches the basal membrane beneath the decidual cells, it penetrates it and further invades into the uterine stroma. Finally, the whole embryo is embedded in the endometrium. Eventually, the syncytiotrophoblast comes into contact with maternal blood and forms chorionic villi. This is the beginning of placentation.
The penetration of the trophoblast to the endometrium is demonstrated through metalloproteinase MMP-2 and MMP-9.[21] Trophoblasts invade the uterus attempting to reach maternal blood supply, for setting up the foundation for fetal blood flow [22]
Extravillous trophoblasts
Extravillous trophoblasts are cells from the invading villi that migrate into the myometrium of the mother’s uterus. These cells remodel the spiral arteries to improve and secure maternal blood flow to the growing embryo. There is also evidence that this process occurs with the uterine veins, stabilizing them to improve drainage of fetal blood and metabolic wastes.[23] Trophoblasts have also been documented to migrate into various tissues in the mother. Due to this trophoblasts have been implicated in a phenomenon known as fetomaternal microchimerism where fetal cells establish cell lines in maternal tissues.[24]
Secretions
The blastocyst secretes factors for a multitude of purposes during invasion. It secretes several autocrine factors, targeting itself and stimulating it to further invade the endometrium.[25] Furthermore, secretions loosen decidual cells from each other, prevent the embryo from being rejected by the mother, trigger the final decidualization and prevent menstruation.
Autocrine
Human chorionic gonadotropin is an autocrine growth factor for the blastocyst.[25] Insulin-like growth factor 2,[25] on the other hand, stimulates the invasiveness of it.
Dislodging
The syncytiotrophoblasts dislodges decidual cells in their way, both by degradation of cell adhesion molecules linking the decidual cells together as well as degradation of the extracellular matrix between them.
Cell adhesion molecules are degraded by syncytiotrophoblast secretion of tumor necrosis factor-alpha. This inhibits the expression of cadherins and beta-catenin.[25] Cadherins are cell adhesion molecules, and beta-catenin helps to anchor them to the cell membrane. Inhibited expression of these molecules thus loosens the connection between decidual cells, permitting the syncytiotrophoblasts and the whole embryo with them to invade into the endometrium.
The extracellular matrix is degraded by serine endopeptidases and metalloproteinases. Examples of such metalloproteinases are collagenases, gelatinases and stromelysins.[25] These collagenases digest Type-I collagen, Type-II collagen, Type-III collagen, Type-VII collagen and Type-X collagen.[25] The gelatinases exist in two forms; one digesting Type-IV collagen and one digesting gelatin.[25]
Immunosuppressive
The embryo differs from the cells of the mother, and would be rejected as a parasite by the immune system of the mother if it did not secrete immunosuppressive agents. Such agents include platelet-activating factor, human chorionic gonadotropin, early pregnancy factor, prostaglandin E2, interleukin-1 alpha, interleukin 6, interferon-alpha, leukemia inhibitory factor and colony-stimulating factor.
Prevention of menstruation
Human chorionic gonadotropin (hCG) not only acts as an immunosuppressive,[25] but also signals to the mother that she is pregnant, preventing luteolysis of the corpus luteum and menstruation by sustaining the function of the corpus luteum.
Other factors
Other factors secreted by the blastocyst are;
- Embryo-derived histamine-releasing factor
- Tissue plasminogen activator as well as its inhibitors
- Estradiol
- β1-integrins
- Fibroblast growth factor
- CYTL1[26]
- Transforming growth factor alpha
- Inhibin
- Preimplantation factor
Uterus receptivity
To enable implantation, the uterus goes through changes in order to be able to receive the conceptus. Receptivity includes changes to endometrial cells in the formation of pinopodes that help to absorb uterine fluid; changes in the thickness of the endometrium and its blood supply development, and the formation of the decidua. Collectively these changes are known as plasma membrane transformation, and bring the blastocyst nearer to the endometrium and immobilize it. During this stage the blastocyst can still be eliminated by being flushed out of the uterus.[27][28]
Successful implantation is co-dependent on the viability of the embryo, and the receptivity of the uterus.[29] A critical involved factor is the developmental synchrony between the embryo and the uterus.[30] The synchrony gives a short period of receptivity known as the window of implantation, and involves much crosstalk between the blastocyst and the endometrium at this stage.[31][29][32] During adhesion this communication is conveyed by receptor-ligand-interactions, both integrin-matrix and proteoglycan ones. Proteoglycan receptors are found on the surface of the decidua, and their counterparts, the proteoglycans, are found around the trophoblast cells of the blastocyst. This ligand-receptor system is also present just at the implantation window.[25] The blastocyst signals to the endometrium to adapt further to its presence, for example by changes in the cytoskeleton of decidual cells. This, in turn, dislodges the decidual cells from their connection to the underlying basal lamina, which enables the blastocyst to perform the succeeding invasion.[25]
Window of implantation
The window of implantation is a limited timeframe for the successful attachment of the blastocyst. [33] In humans uterine receptivity is optimum on days 20-24 of the secretory phase of the menstrual cycle when luteinizing hormone levels are at their peak. [3][34][29] In humans the window of implantation is only available for 24-36 hours.[35]
The endometrial microbiome has been indicated as having an important role in successful implantation in controlling endometrial cell function, and the function of the local immunity system that prevents pathogen growth. This is associated with the secretion of protective substances.[36][37]
Pinopodes
Pinopodes are formed at the beginning of the window of implantation, and are found in many species.[38][33] They are mushroom-like protrusions from the apical cell membrane of uterine epithelial cells.[33] Pinopodes are formed by the swelling of these epithelial cells, and the fusing together of a number of microvilli, to reach a maximum size.[38] They appear between day 19 and day 21 of gestational age, and are fully formed at day 20.[33] This corresponds to a fertilization age of approximately five to seven days, which corresponds well with the time of implantation. Pinopodes only persist for a maximum of two days, and are seen as the ultrastructural markers of receptivity. [38]
Their development is enhanced by progesterone, and inhibited by estrogens. During the window of implantation, cell to cell adhesion is inhibited by MUC1 a cell surface glycoprotein. The most important attribute of pinopodes is the removal of this glycoprotein from the cell surfaces of the uterine epithelial cells.[3] MUC16 has also been shown to disappear from the cell surfaces with the development of the pinopodes. Some studies have reported that pinopodes entrap cilia, which prevents embryo movement, and during implantation allows close contact and adherence of the embryo.[33]
Pinopodes bring uterine fluid and its macromolecules into the cells by the process of endocytosis. This decreases the volume of the uterus, taking the walls closer to the blastocyst floating in it. Thus, the period of active pinopodes might limit the implantation window.[25] Pinopodes continue to absorb fluid, removing most of it during the early stages of implantation.
Predecidualization
The endometrium increases thickness, becomes vascularized and its glands grow to be tortuous and boosted in their secretions. These changes reach their maximum about seven days after ovulation.
Furthermore, the surface of the endometrium produces a kind of rounded cells, which cover the whole area toward the uterine cavity. This happens about 9 to 10 days after ovulation.[25] These cells are called decidual cells, which emphasises that the whole layer of them is shed off in every menstruation if no pregnancy occurs, just as leaves of deciduous trees. The uterine glands, on the other hand, decrease in activity and degenerate around 8 to 9 days[25] after ovulation in absence of pregnancy.
The decidual cells originate from the stromal cells that are always present in the endometrium, and make up a new layer, the decidua. The rest of the endometrium, in addition, expresses differences between the luminal and the basal sides. The luminal cells form the stratum compactum of the endometrium, in contrast to the basalolateral stratum spongiosum, which consists of the rather spongy stromal cells.[25]
Decidualization
Decidualization expands if pregnancy occurs, further developing the uterine glands, the zona compacta and the epithelium of decidual cells lining it. The decidual cells become filled with lipids and glycogen and take the polyhedral shape characteristic of decidual cells. Factors from the blastocyst also trigger the final formation of decidual cells into their proper form. In contrast, some decidual cells in the proximity of the blastocyst degenerate, providing nutrients for it.[25]
Trigger
It is likely that the blastocyst itself makes the main contribution to this additional growing and sustaining of the decidua. An indication of this is that decidualization occurs at a higher degree in conception cycles than in nonconception cycles.[25] Furthermore, similar changes are observed when giving stimuli mimicking the natural invasion of the embryo.[25]
The embryo releases serine proteases which causes the epithelial cell membrane to depolarize and activates the epithelial sodium channel. This triggers a Ca2+ influx and phosphorylation of CREB. Phosphorylation of CREB upregulates the expression of COX2, which leads to the release of prostaglandin E2 (PGE2) from epithelial cells. PGE2 acts on the stroma cells activating cAMP-related pathways in stromal cell leading to decidualization.[39]
Parts of decidua
The decidua can be organized into separate sections, although they have the same composition.
- Decidua basalis – This is the part of the decidua which is located basalolateral to the embryo after implantation.
- Decidua capsularis – Decidua capsularis grows over the embryo on the luminal side, enclosing it into the endometrium. It surrounds the embryo together with decidua basalis.
- Decidua parietalis – All other decidua on the uterine surface belongs to decidua parietalis.
Decidua throughout pregnancy
After implantation the decidua remains, at least through the first trimester.[25] However, its most prominent time is during the early stages of pregnancy, during implantation. Its function as a surrounding tissue is replaced by the definitive placenta. However, some elements of the decidualization remain throughout pregnancy.[25]
The compacta and spongiosa layers are still observable beneath the decidua in pregnancy. The glands of the spongiosa layer continue to secrete during the first trimester, when they degenerate. However, before that disappearance, some glands secrete unequally much. This phenomenon of hypersecretion is called the Arias-Stella phenomenon,[25] after the pathologist Javier Arias-Stella.
Adaptation of secretions
| Proteins, glycoproteins and peptides
secreted by the uterine glands[25] |
| Matrix-associated: |
| Fibronectin |
| Laminin |
| Entactin |
| Type-IV collagen |
| Heparan sulfate |
| Proteoglycan |
| Integrins |
| – |
| Others: |
| Mucins |
| Prolactin |
| IGFBP-1 |
| Glycodelin |
| Endometrial protein 15 |
| Albumin |
| Beta-Lipoprotein |
| Relaxin |
| Fibroblast growth factor 1 |
| Fibroblast growth factor 2 |
| Pappalysin-1 |
| Stress response protein 27 (SRP-27) |
| CA-125 |
| Beta-endorphin |
| Leu-enkephalin |
| Diamine oxidase |
| Tissue plasminogen activator |
| Renin |
| Progesterone-dependent carbonic anhydrase |
| Lactoferrin |
Not only the lining of the uterus transforms, but the secretion from its glands changes. This change is induced by increased levels of progesterone from the corpus luteum. The target of the secretions is the embryoblast, and has several functions on it.
Nourishment
The embryo spends approximately 72 hours in the uterine cavity before implanting. In that time, it cannot receive nourishment directly from the blood of the mother, and must rely on secreted nutrients into the uterine cavity, e.g. iron and fat-soluble vitamins.[25]
Growth and implantation
In addition to nourishment, the endometrium secretes several steroid-dependent proteins, important for growth and implantation. Cholesterol, and steroids are also secreted.[25] Implantation is further facilitated by synthesis of matrix substances, adhesion molecules and surface receptors for the matrix substances.
Clinical significance
Reproduction in humans is not very efficient. Only around 30% of natural conceptions result in successful pregnancies. Of the failed pregnancies around 85% are due to implantation failure.[40] Implantation failure is considered to be caused by inadequate uterine receptivity in two-thirds of cases, and by problems with the embryo itself in the other third.[41] Most IVF procedures fail because of implantation failure accounting for almost half of all pregnancy failures.[40]
Inadequate uterine receptivity may be caused by abnormal cytokine and hormonal signaling as well as epigenetic alterations.[42] Recurrent implantation failure is a cause of female infertility. Therefore, pregnancy rates can be improved by optimizing endometrial receptivity for implantation.[42] Evaluation of implantation markers may help to predict pregnancy outcome and detect occult implantation deficiency.[42] As part of the organ-on-a-chip program, an endometrium-on-a-chip has been developed to model the functioning of the endometrium that could more clearly identify causes of implantation failure.[43] Organoids have also been developed to model the endometrium and its role in implantation.[44]
In women with more than three implantation failures in assisted reproduction, a review of several small randomized controlled studies estimated that the use of adjunct low molecular weight heparin improves live birth rate by approximately 80%.[45] Luteal phase support can include the use of progesterone and human chorionic gonadotropin (hCG) to improve the chances of a successful implantation.[46]
Zinc deficiency
Zinc is crucial in pre-conception, (and successful pregnancy), and its deficiciency can lead to incompetent blastocyst development. Once an egg is fertilized zinc is released in a zinc spark which promotes changes that include the hardening of the zona pellucida preventing polyspermy.[47]
Implantation bleeding
Implantation bleeding is a small amount of light vaginal bleeding or spotting that can occur in early pregnancy due to the fertilized egg implanting in the uterus.[48][49] Bleeding and spotting are common during the luteal phase of the menstrual cycle, and early stages of pregnancy, and are unrelated to implantation. Implantation bleeding occurs between 7 to 14 days after fertilization,[50] and may be accompanied by symptoms such as cramping, nausea, breast tenderness, and headaches.[51] Implantation bleeding can be distinguished from period bleeding by color, clotting, strength and duration of flow.[52][53]
See also
- Embryonic diapause
- Hofbauer cells
- Reichert’s membrane
References
- Niringiyumukiza JD, Cai H, Xiang W (May 2018). "Prostaglandin E2 involvement in mammalian female fertility: ovulation, fertilization, embryo development and early implantation". Reproductive Biology and Endocrinology. 16 (1): 43. doi:10.1186/s12958-018-0359-5. PMC 5928575. PMID 29716588.
- Imakawa K, Bai R, Fujiwara H, Kusama K (January 2016). "Conceptus implantation and placentation: molecules related to epithelial-mesenchymal transition, lymphocyte homing, endogenous retroviruses, and exosomes". Reproductive Medicine and Biology. 15 (1): 1–11. doi:10.1007/s12522-015-0215-7. PMC 5715838. PMID 29259417.
- Kim SM, Kim JS (December 2017). "A Review of Mechanisms of Implantation". Development & Reproduction. 21 (4): 351–359. doi:10.12717/DR.2017.21.4.351. PMC 5769129. PMID 29359200.
- McGowen MR, Erez O, Romero R, Wildman DE (2014). "The evolution of embryo implantation". The International Journal of Developmental Biology. 58 (2–4): 155–161. doi:10.1387/ijdb.140020dw. PMC 6053685. PMID 25023681.
- Moore KL (2020). The developing human: clinically oriented embryology (Eleventh ed.). Edinburgh. pp. 35–42. ISBN 978-0-323-61154-1.
- Cakmak H, Taylor HS (March 2011). "Implantation failure: molecular mechanisms and clinical treatment". Human Reproduction Update. 17 (2): 242–253. doi:10.1093/humupd/dmq037. PMC 3039220. PMID 20729534.
- Ochoa-Bernal MA, Fazleabas AT (March 2020). "Physiologic Events of Embryo Implantation and Decidualization in Human and Non-Human Primates". International Journal of Molecular Sciences. 21 (6): 1973. doi:10.3390/ijms21061973. PMC 7139778. PMID 32183093.
- Gauster M, Moser G, Wernitznig S, Kupper N, Huppertz B (June 2022). "Early human trophoblast development: from morphology to function". Cellular and Molecular Life Sciences. 79 (6): 345. doi:10.1007/s00018-022-04377-0. PMC 9167809. PMID 35661923.
- Sadler TW (2010). Langman's medical embryology (11th ed.). Philadelphia: Lippincott William & Wilkins. p. 45. ISBN 9780781790697.
- Boron 2004, p. 1172.
- Seshagiri, Polani B.; Vani, Venkatappa; Madhulika, Pathak (March 2016). "Cytokines and Blastocyst Hatching". American Journal of Reproductive Immunology. pp. 208–217. doi:10.1111/aji.12464. Retrieved 2 November 2022.
- IVF-infertility.com
- Boron 2004, p. 1173.
- Margarit L, Taylor A, Roberts MH, Hopkins L, Davies C, Brenton AG, et al. (December 2010). "MUC1 as a discriminator between endometrium from fertile and infertile patients with PCOS and endometriosis". The Journal of Clinical Endocrinology and Metabolism. 95 (12): 5320–5329. doi:10.1210/jc.2010-0603. PMID 20826587.
- Carson DD, Julian J, Lessey BA, Prakobphol A, Fisher SJ (September 2006). "MUC1 is a scaffold for selectin ligands in the human uterus". Frontiers in Bioscience. 11 (1): 2903–2908. doi:10.2741/2018. PMID 16720361.
- Francis LW, Yao SN, Powell LC, Griffiths S, Berquand A, Piasecki T, et al. (February 2021). "Highly glycosylated MUC1 mediates high affinity L-selectin binding at the human endometrial surface". Journal of Nanobiotechnology. 19 (1): 50. doi:10.1186/s12951-021-00793-9. PMC 7890821. PMID 33596915.
- Genbacev OD, Prakobphol A, Foulk RA, Krtolica AR, Ilic D, Singer MS, et al. (January 2003). "Trophoblast L-selectin-mediated adhesion at the maternal-fetal interface". Science. 299 (5605): 405–408. Bibcode:2003Sci...299..405G. doi:10.1126/science.1079546. PMID 12532021. S2CID 15462671.
- Singh H, Aplin JD (July 2009). "Adhesion molecules in endometrial epithelium: tissue integrity and embryo implantation". Journal of Anatomy. 215 (1): 3–13. doi:10.1111/j.1469-7580.2008.01034.x. PMC 2714633. PMID 19453302.
- Gipson IK, Blalock T, Tisdale A, Spurr-Michaud S, Allcorn S, Stavreus-Evers A, Gemzell K (January 2008). "MUC16 is lost from the uterodome (pinopode) surface of the receptive human endometrium: in vitro evidence that MUC16 is a barrier to trophoblast adherence". Biology of Reproduction. 78 (1): 134–142. doi:10.1095/biolreprod.106.058347. PMID 17942799. S2CID 44999196.
- Gardiner C, Tannetta DS, Simms CA, Harrison P, Redman CW, Sargent IL (2011). "Syncytiotrophoblast microvesicles released from pre-eclampsia placentae exhibit increased tissue factor activity". PLOS ONE. 6 (10): e26313. Bibcode:2011PLoSO...626313G. doi:10.1371/journal.pone.0026313. PMC 3194796. PMID 22022598.
- Zhu JY, Pang ZJ, Yu YH (2012). "Regulation of trophoblast invasion: the role of matrix metalloproteinases". Reviews in Obstetrics & Gynecology. 5 (3–4): e137–e143. PMC 3594863. PMID 23483768.
- Carter AM, Enders AC, Pijnenborg R (March 2015). "The role of invasive trophoblast in implantation and placentation of primates". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 370 (1663): 20140070. doi:10.1098/rstb.2014.0070. PMC 4305171. PMID 25602074.
- Moser G, Weiss G, Sundl M, Gauster M, Siwetz M, Lang-Olip I, Huppertz B (March 2017). "Extravillous trophoblasts invade more than uterine arteries: evidence for the invasion of uterine veins". Histochemistry and Cell Biology. 147 (3): 353–366. doi:10.1007/s00418-016-1509-5. PMC 5344955. PMID 27774579.
- Dawe GS, Tan XW, Xiao ZC (January 2007). "Cell migration from baby to mother". Cell Adhesion & Migration. 1 (1): 19–27. doi:10.4161/cam.4082. PMC 2633676. PMID 19262088.
- Boron W, Boulpaep E (2004). Medical Physiology: A Cellular And Molecular Approaoch. Oxford: Elsevier. ISBN 1-4160-2328-3. OCLC 61527528.
- Ai Z, Jing W, Fang L (2015). "Cytokine-Like Protein 1(Cytl1): A Potential Molecular Mediator in Embryo Implantation". PLOS ONE. 11 (1): e0147424. doi:10.1371/journal.pone.0147424. PMC 4723121. PMID 26800213.
- Murphy CR (August 2004). "Uterine receptivity and the plasma membrane transformation". Cell Research. 14 (4): 259–267. doi:10.1038/sj.cr.7290227. PMID 15353123. S2CID 24023502.
- Murphy CR, Shaw TJ (December 1994). "Plasma membrane transformation: a common response of uterine epithelial cells during the peri-implantation period". Cell Biology International. 18 (12): 1115–1128. doi:10.1006/cbir.1994.1038. PMID 7703952. S2CID 22769575.
- Yang Y, Zhu QY, Liu JL (November 2021). "Deciphering mouse uterine receptivity for embryo implantation at single-cell resolution". Cell Proliferation. 54 (11): e13128. doi:10.1111/cpr.13128. PMC 8560620. PMID 34558134.
- Teh WT, McBain J, Rogers P (November 2016). "What is the contribution of embryo-endometrial asynchrony to implantation failure?". Journal of Assisted Reproduction and Genetics. 33 (11): 1419–1430. doi:10.1007/s10815-016-0773-6. PMC 5125144. PMID 27480540.
- Sternberg AK, Buck VU, Classen-Linke I, Leube RE (August 2021). "How Mechanical Forces Change the Human Endometrium during the Menstrual Cycle in Preparation for Embryo Implantation". Cells. 10 (8): 2008. doi:10.3390/cells10082008. PMC 8391722. PMID 34440776.
- Liang J, Wang S, Wang Z (November 2017). "Role of microRNAs in embryo implantation". Reprod Biol Endocrinol. 15 (1): 90. doi:10.1186/s12958-017-0309-7. PMC 5699189. PMID 29162091.
- Quinn, KE; Matson, BC; Wetendorf, M; Caron, KM (5 February 2020). "Pinopodes: Recent advancements, current perspectives, and future directions". Molecular and Cellular Endocrinology. 501: 110644. doi:10.1016/j.mce.2019.110644. PMC 6962535. PMID 31738970.
- Xiao Y, Sun X, Yang X, Zhang J, Xue Q, Cai B, Zhou Y (June 2010). "Leukemia inhibitory factor is dysregulated in the endometrium and uterine flushing fluid of patients with adenomyosis during implantation window". Fertility and Sterility. 94 (1): 85–89. doi:10.1016/j.fertnstert.2009.03.012. PMID 19361790.
- Ng SW, Norwitz GA, Pavlicev M, Tilburgs T, Simón C, Norwitz ER (June 2020). "Endometrial Decidualization: The Primary Driver of Pregnancy Health". Int J Mol Sci. 21 (11). doi:10.3390/ijms21114092. PMC 7312091. PMID 32521725.
- Crha I, Ventruba P, Žáková J, Ješeta M, Pilka R, Lousová E, Papíková Z (2019). "Uterine microbiome and endometrial receptivity". Ceska Gynekologie. 84 (1): 49–54. PMID 31213058.
- Moreno I, Codoñer FM, Vilella F, Valbuena D, Martinez-Blanch JF, Jimenez-Almazán J, et al. (December 2016). "Evidence that the endometrial microbiota has an effect on implantation success or failure". American Journal of Obstetrics and Gynecology. 215 (6): 684–703. doi:10.1016/j.ajog.2016.09.075. PMID 27717732.
- Zhao Y, He D, Zeng H, Luo J, Yang S, Chen J, Abdullah RK, Liu N (September 2021). "Expression and significance of miR-30d-5p and SOCS1 in patients with recurrent implantation failure during implantation window". Reprod Biol Endocrinol. 19 (1): 138. doi:10.1186/s12958-021-00820-2. PMC 8425163. PMID 34496883.
- Ruan YC, Guo JH, Liu X, Zhang R, Tsang LL, Dong JD, et al. (July 2012). "Activation of the epithelial Na+ channel triggers prostaglandin E₂ release and production required for embryo implantation". Nature Medicine. 18 (7): 1112–1117. doi:10.1038/nm.2771. PMID 22729284. S2CID 5116022.
- Wang Y, Jiang X, Jia L, Wu X, Wu H, Wang Y, et al. (2022). "A Single-Cell Characterization of Human Post-implantation Embryos Cultured In Vitro Delineates Morphogenesis in Primary Syncytialization". Frontiers in Cell and Developmental Biology. 10: 835445. doi:10.3389/fcell.2022.835445. PMC 9240912. PMID 35784461.
- Melford SE, Taylor AH, Konje JC (2013). "Of mice and (wo)men: factors influencing successful implantation including endocannabinoids". Human Reproduction Update. 20 (3): 415–428. doi:10.1093/humupd/dmt060. PMID 24306146.
- Cakmak H, Taylor HS (2010). "Implantation failure: molecular mechanisms and clinical treatment". Human Reproduction Update. 17 (2): 242–253. doi:10.1093/humupd/dmq037. PMC 3039220. PMID 20729534.
- Ahn J, Yoon MJ, Hong SH, Cha H, Lee D, Koo HS, et al. (September 2021). "Three-dimensional microengineered vascularised endometrium-on-a-chip". Human Reproduction. 36 (10): 2720–2731. doi:10.1093/humrep/deab186. PMC 8450871. PMID 34363466.
- Rawlings TM, Makwana K, Tryfonos M, Lucas ES (July 2021). "Organoids to model the endometrium: implantation and beyond". Reprod Fertil. 2 (3): R85–R101. doi:10.1530/RAF-21-0023. PMC 8801025. PMID 35118399.
- Potdar N, Gelbaya TA, Konje JC, Nardo LG (2013). "Adjunct low-molecular-weight heparin to improve live birth rate after recurrent implantation failure: a systematic review and meta-analysis". Human Reproduction Update. 19 (6): 674–684. doi:10.1093/humupd/dmt032. PMID 23912476.
- Farquhar C, Marjoribanks J (August 2018). "Assisted reproductive technology: an overview of Cochrane Reviews". The Cochrane Database of Systematic Reviews. 2018 (8): CD010537. doi:10.1002/14651858.CD010537.pub5. PMC 6953328. PMID 30117155.
- Garner TB, Hester JM, Carothers A, Diaz FJ (May 2021). "Role of zinc in female reproduction". Biology of Reproduction. 104 (5): 976–994. doi:10.1093/biolre/ioab023. PMC 8599883. PMID 33598687.
- Sheiner E (2011). Bleeding during pregnancy a comprehensive guide. New York: Springer. p. 9. ISBN 978-1-441-99810-1.
- Harville EW, Wilcox AJ, Baird DD, Weinberg CR (September 2003). "Vaginal bleeding in very early pregnancy". Human Reproduction. 18 (9): 1944–1947. doi:10.1093/humrep/deg379. PMID 12923154.
- "Implantation Bleeding vs A Period: How To Tell The Difference". ReUnite Rx. 2020-12-09. Retrieved 2021-07-29.
- Heger, L. Demosthenes E (ed.). "What do implantation cramps feel like? How to tell the difference between implantation and menstrual cramping". Insider. Retrieved 2021-07-29.
- Aggarwal K (2020). "Vaginal Bleeding in Early Pregnancy". In Sharma A (ed.). Labour Room Emergencies. Singapore: Springer. pp. 155–161. doi:10.1007/978-981-10-4953-8_18. ISBN 978-981-10-4953-8. S2CID 212812977.
- "How Do I Know if It's Implantation Bleeding?". MedicineNet. Retrieved 2021-07-29.
Books
- Boron W, Boulpaep E (2004). Medical Physiology: A Cellular And Molecular Approaoch. Oxford: Elsevier. ISBN 1-4160-2328-3. OCLC 61527528.