Asteraceae
Les Asteraceae (Astéracées) constituent une grande famille de plantes dicotylédones, appelées aussi « Composées » (Compositae, nom. cons.)[1],[2] ou plus rarement « Composacées »[1],[2] du fait que ce que l'on prend à première vue pour des « fleurs » chez ces plantes est en réalité des « composés » de fleurs minuscules, réunies en inflorescences appelées « capitules ».
Cet article concerne la famille des Composées en botanique. Pour les autres sens du mot « composée », voir Composition.

| Règne | Plantae |
|---|---|
| Sous-règne | Tracheobionta |
| Division | Magnoliophyta |
| Classe | Magnoliopsida |
| Sous-classe | Asteridae |
| Ordre | Asterales |
Giseke (1792)
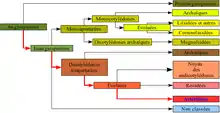
| Clade | Angiospermes |
|---|---|
| Clade | Dicotylédones vraies |
| Clade | Astéridées |
| Clade | Campanulidées |
| Ordre | Asterales |
| Famille | Asteraceae |

Cette famille comprend près de 23 500 espèces réparties en 1 600 genres environ, ce qui en fait la deuxième famille du monde végétal et des plantes à fleurs, derrière les Orchidacées (25 000 espèces) mais devant les Fabacées[3]. Le métabolisme secondaire, l'inflorescence capitulaire et la plasticité écologique sont responsables du succès évolutif de cette famille[4] à répartition cosmopolite (sauf l'Antarctique[Note 1], l'inlandsis du Groenland et l'archipel arctique canadien), mais principalement dans les régions tempérées[5]. Les principaux représentants de cette famille se développent essentiellement dans les régions sujettes à la sécheresse, en dehors de la compétition des arbres des forêts tropicales humides[6].
Ce sont très majoritairement des plantes herbacées, même si la famille comprend aussi des arbres, des arbustes ou des lianes[3].
Étymologie
Le nom vient du genre type Aster, mot latin signifiant étoile, en référence aux capitules étoilés des fleurs.
Compositae est un nom alternatif de cette famille, basé sur les capitules qui sont composés de plusieurs fleurs. Cependant, l’appellation n'étant pas basée sur un nom de genre, on lui a préféré Asteraceae, conformément au code international de nomenclature botanique, Compositae faisant partie des noms dits nomen conservandum, c'est-à-dire « conservés par un long usage »[Note 2].
Caractéristiques générales
Habitus
Les astéracées sont principalement des plantes herbacées, vivaces par des racines formant un pivot simple ou ramifié[7], parfois tubérisées (la molécule de réserve étant l'inuline, polymère du fructose, fibre alimentaire et prébiotique étudié pour son rôle sur le microbiote intestinal)[8]. Certaines sont des plantes annuelles (Helianthus, Tagetes, soucis, laitues). Quelques-unes sont arbustives (Vernonia arborea (en) et Leucomeris (en) au Népal). Certaines sont des xérophytes (Proustia (en), Baccharis), des hydrophytes (Bidens, Cotula) ou hélophytes (Caesulia axillaris (en), Sphaeranthus indicus (en)) se développant dans les rizières. Il existe quelques rares lianes grimpantes (Mikania)[7].
Appareil végétatif

Les feuilles, toujours sans stipules, sont le plus souvent alternes mais parfois opposées (Arnica, Helianthus), rarement verticillées (Eupatoires), ou regroupées en rosette (Bellis, Taraxacum). Dans de nombreux cas, la souche de la rosette basale émet des racines adventives. Les astéracées ont ainsi la capacité de coloniser l'espace à courte distance par reproduction asexuée ou clonale, se développent particulièrement dans les formations végétales à dominante herbacée[9]. Les stratégies de croissance confèrent aux plantes à rosettes un grand pouvoir compétitif et détermine en grande partie la mobilité végétative. Cette mobilité par croissance clonale est également modulée par des facteurs biotiques et abiotique de la plante : compétition pour la lumière au cours des stades précoces de la succession végétale, perturbations (sécheresse récurrente, action anthropique), richesse du sol. La mobilité végétative combinée à l'établissement de nouveaux individus issus de graines par reproduction sexuée expliquerait la dynamique végétale dans un milieu donné[10].
Les feuilles sont souvent simples, mais montrent une grande diversité dans la forme et l'incision (entières à profondément découpées). Dans les hautes montagnes tropicales, elles peuvent devenir succulentes (principalement dans les genres Senecio, Espeletia et Othonna (en)) ou au contraire se réduire à des écailles chez les xérophytes[8].
La tige herbacée est généralement dressée (érigée) mais elle peut aussi être couchée (prostrée) ou plus rarement grimpante. Elle est parfois ligneuse à la base (Armoise, Tanaisie) et transformée en tubercule au goût d'artichaut (Helianthus maximiliani (en), topinambour)[7].
Anabolisme
Les Astéracées présentent soit des canaux résinifères ou oléifères (la plupart des genres, donnant des huiles essentielles terpéniques, comme dans l'Absinthe ou la Camomille), soit des canaux laticifères (tribu des Lactaceae, laitues et laiterons)[11]. Les résines et les latex suintent à la suite de traumatismes (sécheresse, affections fongiques ou bactériennes, attaques d'insectes, blessures mécaniques telles que des incisions), formant en séchant une barrière protectrice contenant notamment des éléments antimicrobiens qui luttent contre la pénétration des pathogènes et contre les herbivores[12].
Les Astéracées produisent des alcaloïdes pyrrolizidiniques, métabolites secondaires également synthétisés par d'autres familles (Boraginacées, Fabacées, Orchidacées…)[13]. Ces métabolites qui illustrent un phénomène de convergence évolutive chimique dans la stratégie de défense des plantes contre les herbivores, ont des effets hépatotoxiques, ce qui explique qu'une consommation inconsidérée de ces plantes peut présenter un danger d'intoxication[13]. Cette famille partage avec les Apiacées la capacité de synthèse de deux nouvelle classes de répulsifs, les lactones sesquiterpéniques[14] et polyacétyléniques (plus de 1 100 dérivés de polyacétylène et de substances biogénétiquement apparentées), qui ont notamment des propriétés cytotoxiques, antimicrobiennes, anti-inflammatoires, neurotoxiques et phototoxiques[15].
Les astéracées stockent leurs réserves énergétiques sous forme de fructanes, principalement l'inuline, plutôt que d'amidon (Métabolisme#Glucides et glycanes). Cette particularité peut être à l'origine de légers désagréments digestifs (ballonements) ou au contraire vue comme un avantage diététique (probiotiques)[8] lors de l'utilisation de ces plantes comme légumes (artichauts, salsifis, scorsonère, topinambour, poire de terre, certaines variétés mexicaines de dahlias…). L'inuline est aussi une matière première de l’industrie alimentaire et potentiellement de l'industrie des plastiques (chicorée industrielle). Étymologiquement, inuline dérive de Inula helenium ou grande aunée, une astéracée dont la racine en est très riche.
Appareil reproducteur


Les Astéracées ont des fleurs minuscules, sessiles (sans pédicelles mais souvent axillées par des bractées internes prenant la forme de paillettes), réunies en inflorescence appelée capitules, c'est-à-dire serrées les unes à côté des autres. Ces capitules élémentaires peuvent à leur tour être rassemblées en grappe, cyme ou souvent corymbe (à l'origine de l'appellation « Corymbifères »). Deux tribus principales sont distinguées selon l'inflorescence : les Cichorioideae à capitules normalement homogames (fleurs toutes stamino-pistillées) constitués de fleurs uniquement ligulées ou toutes tubulées (capitules discoïdes) ; les Asteroideae, capitules polygames (dimorphisme floral : fleurs stamino-pistillées et unisexuées) composés entièrement de fleurs tubulées (capitules disciformes) ou avec en périphérie des fleurs ligulées (capitules radiés). Ils forment une inflorescence surévoluée à laquelle on donne le nom de pseudanthe car chaque capitule simule une fleur entière[16]. Les fleurs sont placées sur l'extrémité d'un rameau ou d'une tige et entourées d'une structure formée par des bractées florales formant une structure en forme de coupe ou de collerette appelée involucre, au rôle protecteur (les bractées charnues de l'artichaut sont d'ailleurs comestibles). Ainsi, contrairement au terme de la langue commune, issu d'un usage très ancien[Note 3], ce qu'on appelle une « fleur » de tournesol, de chardon ou de pissenlit n'est pas « une » fleur mais un pseudanthium (fausse fleur), en l'occurrence un capitule formé d'une multitude de petites fleurs. La disposition des fleurs peut former des spirales régulières dextres et sénestres qui suivent les règles de la suite de Fibonacci[17].
La fleur des Astéracées est très particulière. Elle est pentamère à 5 pétales soudés. Le calice est absent ou réduit (sa fonction protectrice est assurée par l'involucre), les sépales étant en forme de bourrelets, d'écailles ou de soies accrescentes). Les étamines sont également soudées par leurs filets à la base de la corolle et par leurs anthères à déhiscence longitudinale et introrse : c'est un exemple classique de synanthérie, ce qui a valu cette famille le qualificatif de Synanthérées (littéralement anthères soudés)[18]. Néanmoins les anthères ne sont pas concrescentes c'est-à-dire soudées au cours de leur développement, mais simplement cohérentes. La synanthérie permet une présentation à piston du pollen : lorsque le pollinisateur se dirige vers le centre du capitule où les fleurs sont au stade mâle, il stimule le style (muni de poils spécialisés) qui éjecte, tel un piston hors d'un cylindre, les grains de pollen sur l'insecte qui, en visitant d'autres inflorescences, les dépose sur les fleurs femelles du pourtour[19]. La pollinisation est en général entomogame (l'inflorescence attire un large éventail de pollinisateurs généralistes : abeilles, mouches, papillons et scarabées), la protandrie favorisant la fécondation croisée mais si cette fécondation échoue, l'autogamie est possible grâce aux « brosses à pollen » situées sous les stigmates[20]. La croissance rapide du style bifide permet un brossage du pollen et sa récupération. Une fois que le stigmate a traversé le tube formé par les anthères, les stigmates se déplient et exposent leur face gluante au pollen. Un disque nectarifère est à la base du style (nectar souvent protégé de la pluie par des auricules à la base des anthères), au-dessus de l'ovaire infère uniloculaire[21] formé par la soudure de deux carpelles ouverts uniovulés (un seul ovule anatrope). Le style se termine par deux stigmates qui portent une brosse de poils sur leur sommet ou à leur base[20].
Les fruits indéhiscents sont des sortes d'akènes (le terme de cypsèle est plus approprié car la cypsèle résulte de la transformation de deux carpelles, contrairement à l'akène), souvent comprimés, parfois surmontés d'un bec et couronnés d'un pappus sessile ou stipité (aigrette de soies lisses, dentées ou plumeuses), contenant une graines sans albumen, avec des cotylédons volumineux chargés de graisses (certaines espèces ont des graines si riches en huile qu'elles ont été sélectionnées comme oléagineux). Ce pappus est adapté à la zoochorie et à l'anémochorie, notamment à la dispersion transocéanique des graines par le vent, ce qui favorise la spéciation allopatrique[22].
Les pétales d'Astéracées à corolle à « œil de bœuf » (gorge rouge brune) comportent des sillons nanométriques donnant une iridescence. D'autres Astéracées ont des pétales à cellules coniques qui présentent un réseau de crêtes nanométriques à l'origine du même phénomène : ces sillons et crêtes agissent comme un réseau de diffraction qui décompose la lumière blanche (comme le spectre lumineux formé par un prisme ou les irisations à la surface d'un CD) et reflète avant tout les rayons bleus et ultraviolets[23]. Ces Astéracées, comme de nombreuses plantes, n'ont pas la capacité génétique et biochimique de produire des pigments dans le spectre bleu à ultraviolet. Elles créent ainsi cette iridescence afin d'attirer les pollinisateurs grâce à un guide à nectar visible par les insectes mais non par les hommes[24].
Types de capitules des Astéracées
On peut diviser les capitules des Astéracées en quatre groupes :
- les liguliflores (chicorée, pissenlit, laitue, etc.), où le capitule est composé uniquement de fleurs ligulées (parfois appelées demi-fleurons). Celles-ci présentent chacune une languette, ou ligule ; les équivalents des pétales sont soudés, généralement par cinq, parfois par trois, reconnaissables seulement aux dents de la languette, et où un pétale prédomine (fleur irrégulière) ;
- les tubuliflores (chardon, cirse, centaurée, etc.), dont le capitule n'est composé que de fleurs régulières, tubulées (ou fleurs tubulaires parfois appelées fleurons). Elles présentent chacune un tube terminé par des lèvres imperceptibles ou s'ouvrant plus ou moins largement en cinq lobes ;
- les labiatiflores (ligneuses tropicales telles que Mutisia) composées uniquement de fleurs zygomorphes bilabiées ;
- les radiés, aux fleurs périphériques ligulées entourant un disque de fleurs tubulées[25] (marguerite, aster, séneçon, etc.).
 Liguliflore : fleurs toutes ligulées (pissenlit)
Liguliflore : fleurs toutes ligulées (pissenlit) Tubuliflore : fleurs toutes tubulées (artichaut)
Tubuliflore : fleurs toutes tubulées (artichaut) Composée labiatiflore (Mutisia)
Composée labiatiflore (Mutisia) Radié : fleurs ligulées et tubulées (marguerite)
Radié : fleurs ligulées et tubulées (marguerite) Capitule radié (Bidens torta)
Capitule radié (Bidens torta) Super capitule (capitule de capitules) (Edelweiss)
Super capitule (capitule de capitules) (Edelweiss)
Pour identifier la plupart des plantes de cette famille, il est nécessaire de récolter des capitules défleuris, portant des fruits mûrs ou, au moins, déjà bien formés. L'observation des bractées de l'involucre est également très importante.
Particularités
Certaines sont connues pour le caractère allergène de leur pollen (genre Ambroisie).
On peut aussi citer une plante emblématique de la flore montagnarde : l'edelweiss.
Phylogénie et taxinomie
Phylogénie
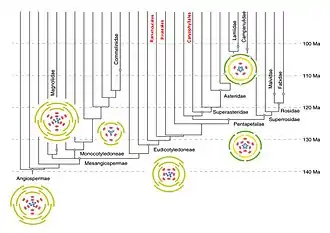
Il pourrait y avoir un ancêtre commun unique à toutes les plantes de cette famille. Il serait originaire d'Amérique du Sud, et daterait de l'Éocène[27].
Ci-dessous : arbre phylogénétique des sous-familles d'Asteraceae.
Le losange désigne un nœud très mal supporté (<50 % de prise en charge du bootstrap), les points des nœuds mal supportés (<80 %).
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Taxinomie
La systématique de cette famille reste délicate au rang des espèces en raison de l'action conjointe de l'apomixie, de l'hybridation et de la polyploïdie[28].
Principaux genres
Les principaux genres sont Senecio (1 500 espèces), Vernonia (1 000 espèces), Cousinia et Eupatorium (600 espèces), Hieracium, Helichrysum et Artemisia (500 espèces), Baccharis et Saussurea (400 espèces), Mikania (300 espèces), Cirsium (270 espèces), Aster et Jurinea (250 espèces), Bidens (200 espèces). La flore française comprend environ 130 genres avec 600 espèces spontanées, auxquelles il faudrait ajouter un certain nombre d'espèces naturalisées. Les principaux genres français sont : Hieracium (les Épervières avec au moins une quarantaine d'espèces), Centaurea (Centaurées avec 36 espèces), Senecio (Séneçons avec 28 espèces), Cirsium et Crepis (Cirses et Crépis avec 25 espèces), Carduus (Chardons avec 19 espèces), Artemisia et Leucanthemum (Armoises et Leucanthèmes avec 17 espèces), Achillea (Achillées avec 13 espèces)[29].
Utilisation
Malgré sa taille importante, cette famille de plantes a donné peu de plantes cultivées ayant une réelle importance économique, relativement moins que des familles plus restreintes comme les Poaceae, les Fabaceae ou les Solanaceae. Aucune espèce d’'Asteraceae ne produit du bois, des fibres ou des matériaux pour la construction. Les principaux produits fournis par cette famille sont des graines oléagineuses, des plantes alimentaires à feuilles, tiges et tubercules, un petit nombre de composés médicinaux, d'insecticides et de plantes aromatiques, ainsi qu'un grand nombre de plantes horticoles ornementales. Certaines espèces sont des mauvaises herbes de l'agriculture[30],[31]. La base de données FAOSTAT ne retient que six produits issus de cette famille : artichaut, carthame, laitue et chicorée, pyrèthre (fleurs séchées) et graines de tournesol[32].
Plantes alimentaires
Sans aucun doute, la principale espèce cultivée de cette famille est la laitue (Lactuca sativa), principale plante consommée comme salade dans le monde. Connue uniquement comme cultigène, c'est vraisemblablement la forme domestiquée de Lactuca serriola. La sélection a conduit à de multiples formes : pommée, batavia, frisée, romaine, iceberg, laitue-asperge, etc.
Les chicorées (genre Cichorium) sont à l'origine de nombreuses salades, dont l'endive (ou chicon), la barbe-de-capucin, la scarole, la frisée, ainsi que la chicorée à café
Le pissenlit (Taraxacum officinale) a les mêmes nombreuses utilisations que la chicorée (salade de pissenlit) mais ne fait pas l'objet d'utilisations industrielles (sauf pour son latex)
L'artichaut et le cardon, variétés de la même espèce (Cynara cardunculus. On consomme le réceptacle floral et la base des bractées de l'artichaut, et le pétiole et le rachis des feuilles du cardon. Leur capitule en fleur ou chardonnette est aussi utilisé comme présure végétale (voir Caillebotte).
Les Asteraceae fournissent aussi des racines et tubercules comestibles : salsifis (Tragopogon porrifolius) et scorsonères (Scorzonera hispanica), topinambour (Helianthus annuus), hélianthi (Helianthus strumosus) et poire de terre (Smallanthus sonchifolius).
Elle fournissent également des graines oléagineuses : graines de tournesol (Helianthus) et de carthame (Carthamus) dont on extrait de l'huile. Le tourteau de tournesol, coproduit de la trituration des graines, est utilisé en alimentation animale. Moins riche en protéines que le tourteau de soja, il a une teneur élevée en acides aminés soufrés, particulièrement en méthionine. On extrait des graines de cardon (Cynara cardunculus var. altilis) une huile de cardon (en) similaire dans sa composition aux précédentes. Les graines de tournesol sont parfois consommées entières, grillées et salées, comme friandises, ou décortiquées comme ingrédient de cuisine[33]. Elles sont aussi appréciées comme aliment pour de nombreuses espèces d'oiseaux de cage[34].
Une espèce sauvage, Gundelia tournefortii, récoltée jeune, est consommée comme légume au Moyen-Orient. La tige, les feuilles (dont les épines sont retirées) et le capitule encore peu développé, sont préparés de diverses manières. La base des jeunes feuilles est considérée comme une gourmandise[35].
 Laitue cultivée (Lactuca sativa).
Laitue cultivée (Lactuca sativa)..jpg.webp) Endives (Cichorium intybus var. foliosum).
Endives (Cichorium intybus var. foliosum). Cardon (Cynara cardunculus var. altilis).
Cardon (Cynara cardunculus var. altilis)._-_geograph.org.uk_-_478257.jpg.webp) Artichaut (Cynara cardunculus var. scolymus).
Artichaut (Cynara cardunculus var. scolymus).
- Boissons
Certaines espèces d'Asteraceae sont à l'origine de boissons de types très différents[36] :
- des infusions, en particulier avec la grande camomille (Tanacetum parthenium), la matricaire camomille (Matricaria recutita), la camomille romaine (Chamaemelum nobile) ;
- la chicorée, substitut du café produit à partir des racines torréfiées de la chicorée à café (Cichorium intybus) ;
- des liqueurs : l'absinthe, produit de la grande absinthe (Artemisia absinthium), et le génépi, produit à partir de plusieurs espèces du genre Artemisia.
- un apéritif, le cynar, à base, entre autres, de feuilles d'artichaut.
- Édulcorants
La stévia (Stevia rebaudiana) contient des glycosides (rébaudiosides) au pouvoir sucrant 200 à 300 fois supérieur à celui du saccharose, sans apporter de calories[37].
L'inuline, présente dans les organes souterrains de nombreuses Asteraceae, est un édulcorant utilisé dans l'industrie agroalimentaire. La principale source est la racine de chicorée à café qui en contient jusqu'à 30 % chez certains cultivars améliorés[36].
- Plantes aromatiques
- Artemisia, genre qui comprend la grande absinthe, la petite absinthe, le génépi blanc, l'estragon, l'aurone
- Calendula, les soucis utilisés comme condiments de salades
- Chamaemelum nobile, la camomille romaine
- Helichrysum italicum parfois appelée herbe à curry
- Matricaria recutita, la camomille allemande
- Tanacetum parthenium, la grande camomille
- Tanacetum vulgare, la tanaisie commune, aromatique mais toxique
- Tagetes patula, l'œillet d'Inde est un classique de la cuisine géorgienne, il est aussi utilisé comme colorant alimentaire, en particulier dans les aliments industriels pour poules pondeuses, afin de donner au jaune d'œuf la teinte souhaitée (chaque région a traditionnellement une préférence pour l'intensité de la coloration du jaune d'œuf entre jaune délavé et orange foncé)
- Santolina chamaecyparissus, la santoline petit-cyprès.
Plantes ornementales
De nombreuses espèces de composées sont utilisées comme plantes ornementales, pour la fleur coupée, ou comme plantes en pot ou plantes à massifs. Plus de 200 genres sont cultivés à cet effet.
Les principales composées ornementales faisant l'objet d'échanges commerciaux au niveau mondial sont les suivantes (principales espèces cultivées) : ageratum (Ageratum houstonianum), aster (Aster spp.), bleuet (Centaurea cyanus), chrysanthème (Chrysanthemum indicum), cosmos (Cosmos sulphureus), dahlia (Dahlia hybrides), gaillarde (Gaillardia pulchella), gerbera (Gerbera hybrides), pâquerette (Bellis perennis), souci (Calendula arvensis, Calendula officinalis , tagète, rose d'Inde, œillet d'Inde (Tagetes erecta, Tagetes patula, Tagetes tenuifolia), tournesol (Helianthus annuus, Helianthus argophyllus, Helianthus debilis, H. decapetalus, H. ×laetiflorus, H. maximilianii, Helianthus ×multiflorus, H. salicifolius), zinnia (Zinnia elegans)[36].
 Chrysanthème 'Seashell'.
Chrysanthème 'Seashell'. Œillet d'Inde (Tagetes erecta).
Œillet d'Inde (Tagetes erecta). Aster × frikartii.
Aster × frikartii. Calendula officinalis.
Calendula officinalis. Dahlia 'Vulkan'.
Dahlia 'Vulkan'.
Autres usages

Certaines espèces fournissent un insecticide à base de pyréthrines. Les Grecs utilisaient l'herbe aux moucherons, sèche, étendue sous le blé pour éloigner les rongeurs et Palladius consigna ces pratiques dans son De re rustica. En compagnonnage, l'armoise est connue pour être un répulsif des rongeurs.
Le gayule (Parthenium argentatum) produit un latex qui est une source alternative de caoutchouc[38].
Mauvaises herbes

De très nombreuses espèces d’Asteraceae sont considérées comme des mauvaises herbes des cultures. On peut citer par exemple les pissenlits (Taraxacum), les laiterons (Sonchus), les cirses (Cirsium), les ambroisies (Ambrosia), les lampourdes (Xanthium)[39].
Dans certains pays, notamment en France et en Belgique, la lutte contre les chardons (échardonnage) est réglementée, voire obligatoire, le terme « chardon » désignant diverses espèces aux feuilles épineuses. La loi vise surtout les cirses, comme le chardon des champs (Cirsium arvense)[40],[41].
L'ambroisie (Ambrosia artemisiifolia) est particulièrement redoutée. Cette plante d'origine nord-américaine s'est répandue dans tous les continents et est considérée comme envahissante. Outre son impact négatif sur les rendements agricoles et sur la biodiversité, son pollen, hautement allergénique, peut provoquer chez les personnes sensibles une rhinite allergique, de la fièvre ou une dermatite[42].
Toxicité
Les Asteraceae renferment relativement peu d'espèces toxiques, qui sont souvent à l'origine d'empoisonnements du bétail. Les principales sont celles qui synthétisent des alcaloïdes pyrrolizidiniques ou des lactones sesquiterpéniques, ainsi que d'autres molécules mono-, di-ou triterpéniques[43]. Les premières appartiennent surtout aux genres Senecio et Eupatorium. On trouve également des pyrrolizidines chez Cineraria, Doronicum, Erechtites, Gynura, Kleinia, Ligularia, Petasites, Tussilago[43]... Parmi les espèces les plus dangereuses figurent le séneçon du Cap (Senecio inaequidens), le séneçon jacobée (Jacobaea vulgaris), le séneçon de Madagascar (Senecio madagascariensis).
Le chardon à glu (Carlina gummifera), espèce méditerranéenne, est mortelle pour l'homme et l'animal du fait de la présence d'un glycoside diterpénoïde, l'atractyloside. Cette molécule est également présente chez la lampourde glouteron (Xanthium strumarium). Cette plante, relativement rare en Europe, est la cause d'intoxications animales mortelles en Amérique du Nord (porcs, moutons, vaches). Les oiseaux semblent insensibles. La dose létale est estimée à 0,3 % du poids corporel[43].
Un cas particulier est celui d'une espèce sud-américaine, Baccharis coridifolia, cause majeure de perte de bétail en Argentine et au Brésil, qui contient des mycotoxines sesquiterpénoïdes (trichothécènes) élaborées par des champignons telluriques du genre Myrothecium[43].
Notes et références
Notes
- Les deux seules plantes à fleurs vivant sur ce continent sont la Sagine antarctique et la Canche antarctique
- Selon le code, les familles sont désignées par le nom d'un de leurs anciens noms génériques avec la désinence -aceæ (art. 21), mais les noms suivants, consacrés par un long usage, font exception à la règle : Compositae (Composées), Cruciferae (Crucifères), Gramineae (Graminées), Guttiferae, Labiatae (Labiacées, labiées ou lamiacées), Leguminosae (Légumineuses), Palmae, Umbelliferae (art. 22).
- Les botanistes eux-mêmes ont longtemps utilisé le terme de « fleur » pour désigner les capitules. Par exemple, Linné décrit en latin les « minore flore » de Chrysanthemum Spec Plant1753 ; Ramatuelle, dans sa communication à l'Académie des Sciences en 1792, décrit la Camomille à grande fleur ou Anthemis grandiflora (actuellement chrysanthème des fleuristes Acad Sc) en distinguant « la fleur terminale de chaque rameau » des « fleurons » et « demi-fleurons ».
Références
- « Composées », dans le Dictionnaire de l'Académie française, sur Centre national de ressources textuelles et lexicales (consulté le 14 mai 2016).
- Définitions lexicographiques et étymologiques de « Composées » dans le Trésor de la langue française informatisé, sur le site du Centre national de ressources textuelles et lexicales (consulté le 14 mai 2016).
- (en) Jeffrey, C. (2007). Compositae: Introduction with key to tribes. In J. J. W. Kadereit, & C. Jeffrey (Eds.), Flowering plants: Eudicots; asterales, Vol. 8, p. 61–87
- (en) Sherwin Carlquist, « Tribal interrelationships and phylogeny of the Asteraceae », Aliso, vol. 8, no 4, , p. 465 (lire en ligne).
- (en) C. Barry Cox, Peter D. Moore, Richard Ladle, Biogeography. An Ecological and Evolutionary Approach, John Wiley & Sons, , p. 42.
- C. Barry Cox, op. cit., p. 43
- (en) Pooja, Angiosperms, Discovery Publishing House, (lire en ligne), p. 309.
- Michel Botineau, Botanique systématique et appliquée des plantes à fleurs, Lavoisier, , p. 1144.
- (en) Lesley Lovett Doust, « Population dynamics and local specialization in a clonal perennial (Ranunculus Repens): I. The dynamics of ramets in contrasting habitats », Journal of Ecology, vol. 69, no 3, , p. 743-755 (DOI 10.2307/2259633).
- (en) Tomáš Herben, František Krahulec, Věra Hadincová & Sylvie Pecháčková, « Is a grassland community composed of coexisting species with low and high spatial mobility ? », Folia Geobotanica & Phytotaxonomica, vol. 29, no 4, , p. 459-468.
- Michel Botineau, Botanique systématique et appliquée des plantes à fleurs, Lavoisier, , p. 1145.
- (en) Jean H. Langenheim, Plant Resins. Chemistry, Evolution, Ecology, and Ethnobotany, Timber Press, (lire en ligne), p. 129-140.
- (en) A. R. Mattocks, Chemistry and Toxicology of Pyrrolizidine Alkaloids, Academic Press, , p. 26
- Frédéric Dupont et Jean-Louis Guignard, Botanique. Les familles de plantes, Elsevier Masson, , p. 266-267.
- (en) D. A. Konovalov, « Polyacetylene Compounds of Plants of the Asteraceae Family (Review) », Pharmaceutical Chemistry Journal, vol. 48, no 9, , p. 613–631.
- Michel Botineau, Botanique systématique et appliquée des plantes à fleurs, Lavoisier, , p. 1149.
- (en) Yafei Zhao, Teng Zhang, Suvi K. Broholm, Sari Tähtiharju, Katriina Mouhu, Victor A. Albert, Teemu H. Teeri, Paula Elomaa, « Evolutionary Co-Option of Floral Meristem Identity Genes for Patterning of the Flower-Like Asteraceae Inflorescence », Plant Physiology, vol. 172, no 1, , p. 284-296 (DOI 10.1104/pp.16.00779)
- Abderrazak Marouf et Joël Reynaud, La botanique de A à Z, Dunod, , p. 293.
- (en) Peter Leins, Claudia Erbar, « Secondary pollen presentation syndromes of the Asterales — a phylogenetic perspective », Botanische Jahrbücher, vol. 127, no 1, , p. 83-103 (DOI 10.1127/0006-8152/2006/0127-0083, lire en ligne)
- Michel Botineau, Botanique systématique et appliquée des plantes à fleurs, Lavoisier, , p. 1150.
- Une seule loge due à la résorption de la cloison entre les deux carpelles.
- (en) Leendert Pijl, Principles of dispersal in higher plants, Springer-Verlag, , p. 91.
- (en) Edwige Moyroud, Tobias Wenzel, Rox Middleton, Paula J. Rudall, Hannah Banks, Alison Reed, Greg Mellers, Patrick Killoran, M. Murphy Westwood, Ullrich Steiner, Silvia Vignolini & Beverley J. Glover, « Disorder in convergent floral nanostructures enhances signalling to bees », Nature, vol. 203, no 550, , p. 469–474 (DOI 10.1038/nature24285)
- (en) K. Yoshida, M. Mori, T. Kondo, « Blue flower color development by anthocyanins: from chemical structure to cell physiology », Nat Prod Rep., vol. 26, no 7, , p. 884-915 (DOI 10.1039/b800165k)
- Bernard Boulard, Dictionnaire de botanique, Paris, Ellipse, , 398 p. (ISBN 978-2-7298-8845-9, BNF 34955567), p. 171
- (en) Susana Magallon, Peter R. Crane and Patrick S. Herendeen, « Phylogenetic pattern, diversity, and diversification of Eudicots », Annals of the Missouri Botanical Garden, vol. 86, no 2, , p. 297-372 (DOI 10.2307/2666180)
- Barreda VD, Palazzesi L, Tellería MC, Eocene Patagonia fossils of the daisy family, Science, 2010;329:1621
- (en) George Wayne Douglas, Elizabeth J. Stephen, The Sunflower Family (Asteraceae) of British Columbia : Astereae, Anthemideae, Eupatorieae and Inuleae, British Columbia Provincial Museum, , p. 382.
- Michel Botineau, Botanique systématique et appliquée des plantes à fleurs, Lavoisier, , p. 1143.
- (en) Beryl B. Simpson, « Economic importance of Compositae », dans Vicki A. Funk, Alfonso Susanna, Tod F. Stuessy, Randall J. Bayer, Systematics, Evolution, and Biogeography of Compositae, Vienne (Autriche), International Association for Plant Taxonomy (IAPT), , 965 p. (ISBN 978-3-9501754-3-1, lire en ligne), p. 45-58.
- (en) Flora of Australia - Asteraceae 1, t. 37, Commonwealth of Australia, (lire en ligne), p. 5-10.
- (en) « FAOSTAT - Cultures », Organisation des Nations unies pour l'alimentation et l'agriculture (FAO) (consulté le ).
- (en) Jane Roche, « Composition de la graine de tournesol (Helianthus annuus L.) sous l’effet conjugué des contraintes agri-environnementales et des potentiels variétaux (thèse) », Institut national polytechnique de Toulouse, (consulté le ).
- « Quelles graines pour quels oiseaux ? », sur Ornithomedia.com (consulté le ).
- (en) E. Vitek, H. Leschner & M. Armağan, « Gundelia tournefortii L. (Compositae)-an approach », Ann. Naturhist. Mus. Wien, vol. 119, , p. 227–233 (lire en ligne).
- (en) Vicki A. Funk, Alfonso Susanna, Tod F. Stuessy, Randall J. Bayer, Systematics, Evolution, and Biogeography of Compositae, Vienne (Autriche), International Association for Plant Taxonomy (IAPT), , 965 p. (ISBN 978-3-9501754-3-1, lire en ligne).
- « Glycosides de stéviol (E960) », sur Edulcorants.eu (consulté le ).
- « Une étape vers la bioraffinerie du guayule : l’analyse rapide des teneurs en caoutchouc et en résines par spectroscopie », sur CIRAD, (consulté le ).
- (en) « Asteraceae » (consulté le ).
- « Les méthodes de lutte contre les chardons », sur Fourrages Mieux (consulté le ).
- « Fiche n° 67- Lutte contre les chardons : Réglementation », sur Terre saine Poitou-Charentes (consulté le ).
- (en) « Ambrosia artemisiifolia (common ragweed) », sur Invasive Species Compendium (ISC), CABI (consulté le ).
- J. Bruneton, Plantes toxiques, végétaux dangereux pour l’Homme et les animaux, Paris/Cachan, Tec & Doc Lavoisier, , 3e éd., 618 p. (ISBN 2-7430-0806-7), p. 183-208.
Voir aussi
Articles connexes
Bibliographie
- Jose L. Panero, Bonnie S. Crozier, « Macroevolutionary dynamics in the early diversification of Asteraceae », Molecular Phylogenetics and Evolution, vol. 99, , p. 116-132 (DOI 10.1016/j.ympev.2016.03.007, lire en ligne)̩.
- (en) Vicki A. Funk, Alfonso Susanna, Tod F. Stuessy, Randall J. Bayer, Systematics, Evolution, and Biogeography of Compositae, Vienne (Autriche), International Association for Plant Taxonomy (IAPT), , 965 p. (ISBN 978-3-9501754-3-1, lire en ligne).
- (en) Flora of Australia - Asteraceae 1, t. 37, Commonwealth of Australia, (lire en ligne).
- (en) Marta Denisow-Pietrzyk, Łukasz Pietrzyk & Bożena Denisow, « Asteraceae species as potential environmental factors of allergy », Environ Sci Pollut Res Int., vol. 26, no 7, , p. 6290–6300 (DOI 10.1007/s11356-019-04146-w, lire en ligne).
Liens externes
Références taxinomiques
- (en) Référence Tree of Life Web Project : Asteraceae
- (fr) Référence Belles fleurs de France : Asteraceae
- (fr) Référence Belles fleurs de France 2 : Asteraceae
- (en) Référence Flora of North America : Asteraceae
- (en) Référence Madagascar Catalogue : Asteraceae
- (en) Référence Flora of Chile : Asteraceae
- (en) Référence Angiosperm Phylogeny Website : Asteraceae ()
- (en) Référence Jardin botanique du Missouri : Asteraceae (images de Madagascar)
- (en) Référence FloraBase (Australie-Occidentale) : classification Asteraceae
- (en) Référence DELTA Angio : Compositae
- (fr+en) Référence ITIS : Asteraceae
- (en) Référence NCBI : Asteraceae (taxons inclus)
- (fr) Référence Tela Botanica (France métro) : Asteraceae
- (en) Référence GRIN : famille Asteraceae Bercht. & J. Presl (+liste des genres contenant des synonymes)
Autres
- (en) Asteraceae, site du jardin botanique du Missouri
- (en) Nádia Roque, David J. Keil & Alfonso Susanna, « Illustrated glossary of Compositae », sur Conference: Systematics, Evolution and Biogeography of Compositae - Volume: Appendix A, (consulté le ).
- (en) « Global Compositae Database », sur The International Compositae Alliance (TICA) (consulté le ).
 Portail de la botanique
Portail de la botanique