Indiana vesiculovirus
| Indiana vesiculovirus | |
|---|---|
_EM_18_lores.jpg.webp) | |
| TEM micrograph of Indiana vesiculovirus particles | |
| Virus classification | |
| (unranked): | Virus |
| Realm: | Riboviria |
| Kingdom: | Orthornavirae |
| Phylum: | Negarnaviricota |
| Class: | Monjiviricetes |
| Order: | Mononegavirales |
| Family: | Rhabdoviridae |
| Genus: | Vesiculovirus |
| Species: | Indiana vesiculovirus |
| Synonyms[1] | |
| |
Indiana vesiculovirus, formerly Vesicular stomatitis Indiana virus (VSIV or VSV) is a virus in the family Rhabdoviridae; the well-known Rabies lyssavirus belongs to the same family. VSIV can infect insects, cattle, horses and pigs. It has particular importance to farmers in certain regions of the world where it infects cattle. This is because its clinical presentation is identical to the very important foot and mouth disease virus.[2]
The virus is zoonotic and leads to a flu-like illness in infected humans.
It is also a common laboratory virus used to study the properties of viruses in the family Rhabdoviridae, as well as to study viral evolution.[3]
Properties
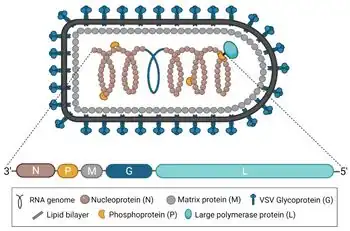
Indiana vesiculovirus is the prototypic member of the genus Vesiculovirus of the family Rhabdoviridae. VSIV is an arbovirus, and its replication occurs in the cytoplasm. Natural VSIV infections encompass two steps, cytolytic infections of mammalian hosts and transmission by insects. In insects, infections are noncytolytic persistent. One confirmed vector of the virus is the phlebotomine sand fly Lutzomyia shannoni.[4] The genome of VSIV is on a single molecule of negative-sense RNA that has 11,161 nucleotides in length,[5] that encodes five major proteins: G protein (G), large protein (L), phosphoprotein (P), matrix protein (M) and nucleoprotein (N):
The VSIV G protein, aka VSVG, enables viral entry. It mediates viral attachment to an LDL receptor (LDLR) or an LDLR family member present on the host cell.[6] Following binding, the VSIV-LDLR complex is rapidly endocytosed. It then mediates fusion of the viral envelope with the endosomal membrane. VSIV enters the cell through partially clathrin-coated vesicles; virus-containing vesicles contain more clathrin and clathrin adaptor than conventional vesicles. Virus-containing vesicles recruit components of the actin machinery for their interaction, thus inducing its own uptake. VSIV G does not follow the same path as most vesicles because transport of the G protein from the ER to the plasma membrane is interrupted by incubation at 15 °C. Under this condition, the molecules accumulate in both the ER and a subcellular vesicle fraction of low density called the lipid-rich vesicle fraction. The material in the lipid-rich vesicle fraction appears to be a post-ER intermediate in the transport process to the plasma membrane (PM). After infection, the VSIV G gene is expressed and is commonly studied as a model for N-linked glycosylation in the endoplasmic reticulum (ER). It is translated into the rough ER where the Glc3-Man9-GlcNac2 oligosaccharide is added by a dolichol-containing protein, to an NXS motif on VSIV G. Sugars are removed gradually as the protein travels to the Golgi apparatus, and it becomes resistant to endoglycosidase H.[7] When synthesized in polarized epithelial cells, the envelope glycoprotein VSV G is targeted to the basolateral PM. VSVG is also a common coat protein for lentiviral vector expression systems used to introduce genetic material into in vitro systems or animal models, mainly because of its extremely broad tropism.
The VSIV L protein is encoded by half the genome, and combines with the phosphoprotein to catalyze replication of the mRNA.
The VSIV M protein is encoded by an mRNA that is 831 nucleotides long and translates to a 229 amino acid-protein. The predicted M protein sequence does not contain any long hydrophobic or nonpolar domains that might promote membrane association. The protein is rich in basic amino acids and contains a highly basic amino terminal domain.
The VSV N protein is required to initiate genome synthesis.[8][9]
Vesicular stomatitis
Signs and diagnosis
The main sign in animals is oral disease appearing as mucosal vesicles and ulcers in the mouth, but also on the udder and around the coronary band. Animals may show systemic signs such as anorexia, lethargy and pyrexia (fever). Disease usually resolves within two weeks, and animals usually recover completely.[2]
Cases of human infection with vesicular stomatitis virus have been described. Most of these cases have been among laboratory workers, veterinarians, and livestock handlers. In most cases, VSV infection has resulted in a short 3 to 5 day illness characterized by fever, headache, myalgia, weakness and occasionally vesicular lesions of the mouth.[10] Serological testing is most commonly performed with an ELISA or complement fixation, and viral isolation can also be attempted.[2]
Treatment and control
No specific treatment is available, but some animals may require supportive fluids or antibiotics for secondary infections.[2]
Control relies on biosecurity protocols, quarantine, isolation and disinfection to ensure the viral disease does not enter a country or herd.[2]
Medical applications
Oncolytic therapy
In healthy human cells the virus cannot reproduce, likely because of the interferon response, which allows the cells to adequately respond to viral infection. The same cannot be said of interferon non-responsive cancer cells, a quality which allows VSIV to grow and lyse oncogenic cells preferentially.[11]
Recently, attenuated VSIV with a mutation in its M protein has been found to have oncolytic properties. Research is ongoing, and has shown VSIV to reduce tumor size and spread in melanoma, lung cancer, colon cancer and certain brain tumors in laboratory models of cancer.[12]
Anti-HIV therapy
VSIV was modified to attack HIV-infected T-cells. The modified virus was called a "trojan horse" virus
Ebola vaccine
Recombinant VSIV has undergone phase 1 trials as a vaccine for Ebola virus.[13]
Recombinant VSIV expressing the Ebola virus glycoprotein has undergone phase III trials in Africa as a vaccine for Ebola virus disease. The vaccine was shown to be 76-100% effective in preventing Ebola virus disease.[14][15] (see also rVSV-ZEBOV vaccine) In December 2019, Merck & Co.'s rVSV-ZEBOV vaccine Ervebo was approved by the Food and Drug Administration to treat individuals 18 and older.[16]
Other Uses
Replication competent rVSV has also been created expressing proteins of Lassa fever and Marburg virus.[17]
Other applications
The VSIV G protein is routinely used in biomedical research to pseudotype retroviral and lentiviral vectors, conferring the ability to transduce a broad range of mammalian cell types with genes of interest.[18]
The VSIV G protein has also been used in cytological studies of trafficking in the endomembrane system. Immunoelectron microscopy suggests that VSIV G protein moves from cis to trans Golgi bodies without being transported between them in vesicles, supporting the cisternal maturation model of Golgi trafficking.[19]
VSV is often used to perform quantitative and computational studies on viral genome replication and transcription.[8][20] Such studies help build a better understanding of viral behavior in the presence and absence of innate immune response.
In 2020, a possible vaccine against COVID-19, the disease caused by SARS-CoV-2, was developed based on modified VSV. The modification involved replacing the VSV's surface protein genes with those for SARS-CoV-2's spike proteins.[21][22]
See also
- Vesiculovirus matrix proteins
- Viral neuronal tracing
- rVSV-ZEBOV vaccine
References
- ↑ "ICTV Taxonomy history: Indiana vesiculovirus" (html). International Committee on Taxonomy of Viruses (ICTV). Archived from the original on 1 July 2023. Retrieved 6 February 2019.
- 1 2 3 4 5 "Vesicular Stomatitis Virus". Archived from the original on 2013-02-22. Retrieved 2023-05-19. reviewed and published by WikiVet, accessed 12 October 2011.
- ↑ Norkin LC (2010). Virology: Molecular Biology and Pathogenesis. Washington DC: American Society for Microbiology Press. ISBN 978-1-55581-453-3.
- ↑ Mann RS, Kaufman PE, Butler JF (2009). "A Sand Fly, Lutzomyia shannoni Dyar (Insecta: Diptera: Psychodidae: Phlebotomine)". EENY-421. Entomology and Nematology. Florida Cooperative Extension Service. University of Florida IFAS. Archived from the original on 2016-09-12. Retrieved 2023-05-19.
- ↑ "VSV complete genome". Archived from the original on 21 August 2018. Retrieved 30 May 2013.
- ↑ Finkelshtein D, Werman A, Novick D, Barak S, Rubinstein M. LDL receptor and its family members serve as the cellular receptors for vesicular stomatitis virus. Proc Natl Acad Sci U S A. 2013 Apr 30;110(18):7306–11.
- ↑ Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P (2002). "Transport from the ER through the Golgi Apparatus". Molecular Biology of the Cell (Fourth ed.). New York: Garland Science. Archived from the original on 2023-03-06. Retrieved 2023-05-19.
- 1 2 Timm C, Gupta A, Yin J (August 2015). "Robust kinetics of an RNA virus: Transcription rates are set by genome levels". Biotechnology and Bioengineering. 112 (8): 1655–62. doi:10.1002/bit.25578. PMC 5653219. PMID 25726926.
- ↑ Davis NL, Wertz GW (March 1982). "Synthesis of vesicular stomatitis virus negative-strand RNA in vitro: dependence on viral protein synthesis". Journal of Virology. 41 (3): 821–32. doi:10.1128/JVI.41.3.821-832.1982. PMC 256819. PMID 6284973.
- ↑ Quiroz E, Moreno N, Peralta PH, Tesh RB. A human case of encephalitis associated with vesicular stomatitis virus (Indiana serotype) infection. Am J Trop Med Hyg. 1988;39(3):312–314. doi:10.4269/ajtmh.1988.39.312
- ↑ Stojdl DF, Lichty B, Knowles S, Marius R, Atkins H, Sonenberg N, Bell JC (July 2000). "Exploiting tumor-specific defects in the interferon pathway with a previously unknown oncolytic virus". Nature Medicine. 6 (7): 821–5. doi:10.1038/77558. PMID 10888934. S2CID 8492631.
- ↑ Ozduman K, Wollmann G, Piepmeier JM, van den Pol AN (February 2008). "Systemic vesicular stomatitis virus selectively destroys multifocal glioma and metastatic carcinoma in brain". The Journal of Neuroscience. 28 (8): 1882–93. doi:10.1523/JNEUROSCI.4905-07.2008. PMC 6671450. PMID 18287505.
- ↑ Agnandji ST, Huttner A, Zinser ME, Njuguna P, Dahlke C, Fernandes JF, et al. (April 2016). "Phase 1 Trials of rVSV Ebola Vaccine in Africa and Europe". The New England Journal of Medicine. 374 (17): 1647–60. doi:10.1056/NEJMoa1502924. PMC 5490784. PMID 25830326.
- ↑ Henao-Restrepo AM, Longini IM, Egger M, Dean NE, Edmunds WJ, Camacho A, et al. (August 2015). "Efficacy and effectiveness of an rVSV-vectored vaccine expressing Ebola surface glycoprotein: interim results from the Guinea ring vaccination cluster-randomised trial". Lancet. 386 (9996): 857–66. doi:10.1016/S0140-6736(15)61117-5. hdl:10144/575218. PMID 26248676. S2CID 40830730.
- ↑ Henao-Restrepo AM, Camacho A, Longini IM, Watson CH, Edmunds WJ, Egger M, et al. (February 2017). "Efficacy and effectiveness of an rVSV-vectored vaccine in preventing Ebola virus disease: final results from the Guinea ring vaccination, open-label, cluster-randomised trial (Ebola Ça Suffit!)". Lancet. 389 (10068): 505–518. doi:10.1016/S0140-6736(16)32621-6. PMC 5364328. PMID 28017403.
- ↑ Commissioner, Office of the (2020-03-24). "First FDA-approved vaccine for the prevention of Ebola virus disease, marking a critical milestone in public health preparedness and response". FDA. Archived from the original on 2019-12-20. Retrieved 2021-10-20.
- ↑ Garbutt M, Liebscher R, Wahl-Jensen V, Jones S, Möller P, Wagner R, et al. (May 2004). "Properties of replication-competent vesicular stomatitis virus vectors expressing glycoproteins of filoviruses and arenaviruses". Journal of Virology. 78 (10): 5458–65. doi:10.1128/jvi.78.10.5458-5465.2004. PMC 400370. PMID 15113924.
- ↑ Cronin J, Zhang XY, Reiser J (August 2005). "Altering the tropism of lentiviral vectors through pseudotyping". Current Gene Therapy. 5 (4): 387–98. doi:10.2174/1566523054546224. PMC 1368960. PMID 16101513.
- ↑ Mironov AA, Beznoussenko GV, Nicoziani P, Martella O, Trucco A, Kweon HS, et al. (December 2001). "Small cargo proteins and large aggregates can traverse the Golgi by a common mechanism without leaving the lumen of cisternae". The Journal of Cell Biology. 155 (7): 1225–38. doi:10.1083/jcb.200108073. PMC 2199327. PMID 11756473.
- ↑ Lim KI, Lang T, Lam V, Yin J (September 2006). "Model-based design of growth-attenuated viruses". PLOS Computational Biology. 2 (9): e116. Bibcode:2006PLSCB...2..116L. doi:10.1371/journal.pcbi.0020116. PMC 1557587. PMID 16948530.
- ↑ SciTechDaily: Researchers Created a Virus That Mimics SARS-CoV-2, the COVID-19 Coronavirus – Here’s Why Archived 2023-02-03 at the Wayback Machine. Source: Washington University School of Medicine. August 19, 2020
- ↑ James Brett Case, Paul W. Rothlauf, Rita E. Chen, Zhuoming Liu, Haiyan Zhao, Arthur S. Kim, Louis-Marie Bloyet, Qiru Zeng, Stephen Tahan, Lindsay Droit, Ma. Xenia G. Ilagan, Michael A. Tartell, Gaya Amarasinghe, Jeffrey P. Henderson, Shane Miersch, Mart Ustav, Sachdev Sidhu, Herbert W. Virgin, David Wang, Siyuan Ding, Davide Corti, Elitza S. Theel, Daved H. Fremont, Michael S. Diamond and Sean P. J. Whelan: Neutralizing Antibody and Soluble ACE2 Inhibition of a Replication-Competent VSV-SARS-CoV-2 and a Clinical Isolate of SARS-CoV-2, Cell Host and Microbe, 1 July 2020, doi:10.1016/j.chom.2020.06.021
External links
- ViralZone: Vesiculovirus Archived 2010-06-20 at the Wayback Machine
- Vesicular Stomatitis Virus Archived 2005-11-23 at the Wayback Machine from The Lab-On-Site Project.
- Disease card Archived 2022-01-20 at the Wayback Machine on World Organisation for Animal Health