Eukaryotic translation elongation factor 1 alpha 1
Elongation factor 1-alpha 1 (eEF1a1) is a translation elongation protein, expressed across eukaryotes. In humans, it is encoded by the EEF1A1 gene.[5][6]



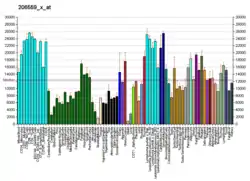
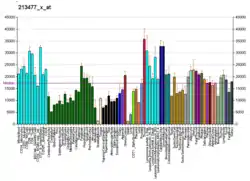
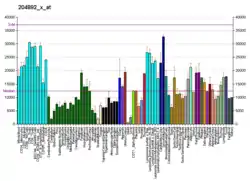
This gene encodes an isoform of the alpha subunit of the elongation factor-1 complex, which is responsible for the enzymatic delivery of aminoacyl tRNAs to the ribosome. This isoform (alpha 1) is expressed in brain, placenta, lung, liver, kidney, and pancreas, and the other isoform (alpha 2) is expressed in brain, heart and skeletal muscle. This isoform is identified as an autoantigen in 66% of patients with Felty's syndrome. This gene has been found to have multiple copies on many chromosomes, some of which, if not all, represent different pseudogenes.[7]
Structure
Mammalian eEF1A possesses two paralogs, eEF1A1 and eEF1A2, with high amino acid sequence homology (approximately 90% identity).[8][9][10][11] The sequences of their promoter regions are also highly similar, though that of the eEF1A2 gene contains an additional 81 bp SV40 small antigen sequence at the 5′-end.[12] The EEF1A1 5' UTR also contains a terminal oligopyrimidine tract.[13] Thus, these two isoforms demonstrate differences in expression and function: eEF1A1 is expressed in most cells while eEF1A2 is only expressed in adult neuronal and muscle cells, and only eEF1A1 induces HSP70 during heat shock.[11]
Function
The eEF1A1 protein is an isoform of the eEF-1 complex alpha subunit, an elongation factor protein, a GTPase, and an actin bundling protein.[7][8][9][14] As an elongation factor, it is known to mediate the recruitment of aminoacyl-tRNA to the A-site of the 80S ribosome during protein synthesis.[8][14][15][16] As a result, this protein is ubiquitously expressed.[8][10][14]
In addition to its role in translation, eEF1A has been shown to play a central role in the nuclear export of proteins.[17][18][19] Thus, eEF1A can be found in both the cytoplasm for translation and in the nucleus for nuclear transport.[11] VHL, PABP1 and other proteins containing a TD-NEM (Transcription Dependent Nuclear Export Motif) are exported by eEF1A in a manner that is dependent on ongoing RNA polymerase II (RNA PolII)-dependent transcription.[17]
Moreover, it participates in several processes required for cell growth and proliferation, including cytoskeleton organization, mitotic apparatus formation, and signal transduction.[8][9][14][19] This protein colocalizes with filamentous actin (F-actin) and is just as abundantly expressed.[9][14] eEF1A putatively binds actin and microtubules at synapses to modulate the cytoskeleton. In neurons, this ability allows for regulation of the number and size of inhibitory postsynaptic complexes like postsynaptic gephyrin clusters.[8] eEF1A has also been found to bind to several kinases, phospholipases, and synaptic proteins.[8] For instance, while associated with actin, it can activate phosphatidylinositol 4-kinase, which then regulates phosphatidylinositol 4-phosphate and phosphatidylinositol 4,5-bisphosphate levels.[14] This protein can also act as a membrane receptor for the cryptic antiadhesive site of fibronectin, thus inhibiting cell anchorage and promoting apoptosis, or anoikis. Though it has not been observed to localize to the cell membrane, it can be found in the outer cell surface.[16] Its role in apoptosis may also contribute to regulation of cell growth and the immune response.[14]
Additional functions of eEF1A1 include: serving as a coactivator of the mineralocorticoid receptors in the heart and kidney to enhance expression of endogenous GILZ, SGK1, and CNKSR3;[10] mediating the TNFα-induced decrease in endothelial nitric oxide synthase mRNA stability;[10] detecting misfolded proteins and targeting them to the proteosome for proteolytic degradation;[19] stabilizing viral and cellular RNAs by binding the 3' region;[11] regulating transcription by recruiting and activating HSF1;[11] and induction of HSP70 during heat shock.[11]
Clinical Significance
Upregulation of eEF1A has been reported in breast cancer samples.[20] Interestingly, however, this upregulation only occurs at the protein level, because the mRNA level is significantly reduced in breast cancer samples.[20] This paradox has been explained by cell cycle-regulated EEF1A1 mRNA expression and stress-induced increase in eEF1A protein levels in breast cancer cells.[20][11] Though its role in metastasis remains unclear, the role of eEF1A in cytoskeleton organization may promote tumor cell motility and thus spread.[9] Alternatively, apoptotic cells may secrete antigens, including eEF1A and other elongation factors, to induce an autoimmune response during cancer. It is postulated that high expression and secretion of elongation factors from tumor tissues, combined with altered levels of eEF1A-derived bacterial peptides in neoplastic disease, may lead to autoimmunity in breast cancer.[14]
As with breast cancer, upregulation of eEF1A expression is associated with prostate cancer and worsened metastasis-free and overall patient survival.[9] Moreover, a truncated form of the eEF1A1 protein, prostate tumour inducing gene 1 (PTI-1), has been detected in prostate carcinoma patient-derived blood samples. As eEF1A1 is over-expressed in osteoblasts, which proliferate and differentiate in the presence of tumor cells, it may serve as a serum biomarker to track the metastatic progression of prostate cancer.[15]
In the case of acute T lymphocytic leukemia, knocking down the eEF1A1 gene produces inhibited proliferation and induced apoptosis of Jurkat cells. These effects may be attributed to the resulting down-regulation of the PI3K/Akt/NF-κB and PI3K/Akt/mTOR signaling pathways.[21]
eEF1A1 is downregulated in myocardial infarction and potentially participates in cardiovascular disease through its interactions with mineralocorticoid receptors in the heart.[10] Its role in heat shock response presents it as a target for treating related diseases like spinal and bulbar muscular atrophy (SBMA) and amyotrophic lateral sclerosis (ALS).[11]
Interactions
Eukaryotic translation elongation factor 1 alpha 1 has been shown to interact with:
References
- GRCh38: Ensembl release 89: ENSG00000156508 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000037742 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Lund A, Knudsen SM, Vissing H, Clark B, Tommerup N (Sep 1996). "Assignment of human elongation factor 1alpha genes: EEF1A maps to chromosome 6q14 and EEF1A2 to 20q13.3". Genomics. 36 (2): 359–61. doi:10.1006/geno.1996.0475. PMID 8812466.
- Wende H, Volz A, Ziegler A (Jul 2000). "Extensive gene duplications and a large inversion characterize the human leukocyte receptor cluster". Immunogenetics. 51 (8–9): 703–13. doi:10.1007/s002510000187. PMID 10941842. S2CID 20719684.
- "Entrez Gene: EEF1A1 eukaryotic translation elongation factor 1 alpha 1".
- Becker M, Kuhse J, Kirsch J (Dec 2013). "Effects of two elongation factor 1A isoforms on the formation of gephyrin clusters at inhibitory synapses in hippocampal neurons". Histochemistry and Cell Biology. 140 (6): 603–9. doi:10.1007/s00418-013-1122-9. PMID 23839781. S2CID 12836593.
- Liu H, Ding J, Chen F, Fan B, Gao N, Yang Z, Qi L (Nov 2010). "Increased expression of elongation factor-1α is significantly correlated with poor prognosis of human prostate cancer". Scandinavian Journal of Urology and Nephrology. 44 (5): 277–83. doi:10.3109/00365599.2010.492787. PMID 20545466. S2CID 13579193.
- Yang J, Fuller PJ, Morgan J, Shibata H, McDonnell DP, Clyne CD, Young MJ (Sep 2014). "Use of phage display to identify novel mineralocorticoid receptor-interacting proteins". Molecular Endocrinology. 28 (9): 1571–84. doi:10.1210/me.2014-1101. PMC 5414797. PMID 25000480.
- Vera M, Pani B, Griffiths LA, Muchardt C, Abbott CM, Singer RH, Nudler E (16 September 2014). "The translation elongation factor eEF1A1 couples transcription to translation during heat shock response". eLife. 3: e03164. doi:10.7554/eLife.03164. PMC 4164936. PMID 25233275.
- Zheng C, Baum BJ (2014). "All human EF1α promoters are not equal: markedly affect gene expression in constructs from different sources". International Journal of Medical Sciences. 11 (5): 404–8. doi:10.7150/ijms.8033. PMC 3970091. PMID 24688302.
- Zhu J, Hayakawa A, Kakegawa T, Kaspar RL (Oct 2001). "Binding of the La autoantigen to the 5' untranslated region of a chimeric human translation elongation factor 1A reporter mRNA inhibits translation in vitro". Biochimica et Biophysica Acta. 1521 (1–3): 19–29. doi:10.1016/s0167-4781(01)00277-9. PMID 11690632.
- Hamrita B, Nasr HB, Hammann P, Kuhn L, Guillier CL, Chaieb A, Khairi H, Chahed K (Sep 2011). "An elongation factor-like protein (EF-Tu) elicits a humoral response in infiltrating ductal breast carcinomas: an immunoproteomics investigation". Clinical Biochemistry. 44 (13): 1097–104. doi:10.1016/j.clinbiochem.2011.06.005. PMID 21704614.
- Rehman I, Evans CA, Glen A, Cross SS, Eaton CL, Down J, Pesce G, Phillips JT, Yen OS, Thalmann GN, Wright PC, Hamdy FC (2012). "iTRAQ identification of candidate serum biomarkers associated with metastatic progression of human prostate cancer". PLOS ONE. 7 (2): e30885. Bibcode:2012PLoSO...730885R. doi:10.1371/journal.pone.0030885. PMC 3280251. PMID 22355332.
- Itagaki K, Naito T, Iwakiri R, Haga M, Miura S, Saito Y, Owaki T, Kamiya S, Iyoda T, Yajima H, Iwashita S, Ejiri S, Fukai F (May 2012). "Eukaryotic translation elongation factor 1A induces anoikis by triggering cell detachment". The Journal of Biological Chemistry. 287 (19): 16037–46. doi:10.1074/jbc.M111.308122. PMC 3346144. PMID 22399298.
- Lee S, Neumann M, Stearman R, Stauber R, Pause A, Pavlakis GN, Klausner RD (Feb 1999). "Transcription-dependent nuclear-cytoplasmic trafficking is required for the function of the von Hippel-Lindau tumor suppressor protein". Molecular and Cellular Biology. 19 (2): 1486–97. doi:10.1128/mcb.19.2.1486. PMC 116077. PMID 9891082.
- Khacho M, Mekhail K, Pilon-Larose K, Pause A, Côté J, Lee S (Dec 2008). "eEF1A is a novel component of the mammalian nuclear protein export machinery". Molecular Biology of the Cell. 19 (12): 5296–308. doi:10.1091/mbc.E08-06-0562. PMC 2592675. PMID 18799616.
- Morrissey C, Schwefel D, Ennis-Adeniran V, Taylor IA, Crow YJ, Webb M (Feb 2015). "The eukaryotic elongation factor eEF1A1 interacts with SAMHD1". The Biochemical Journal. 466 (1): 69–76. doi:10.1042/BJ20140203. PMID 25423367.
- Lin CY, Beattie A, Baradaran B, Dray E, Duijf PH (September 2018). "Contradictory mRNA and protein misexpression of EEF1A1 in ductal breast carcinoma due to cell cycle regulation and cellular stress". Scientific Reports. 8 (1): 13904. Bibcode:2018NatSR...813904L. doi:10.1038/s41598-018-32272-x. PMC 6141510. PMID 30224719.
- Huang Y, Hu JD, Qi YL, Wu YA, Zheng J, Chen YY, Huang XL (Aug 2012). "[Effect of knocking down eEF1A1 gene on proliferation and apoptosis in Jurkat cells and its mechanisms]". Zhongguo Shi Yan Xue Ye Xue Za Zhi. 20 (4): 835–41. PMID 22931638.
- Chang JS, Seok H, Kwon TK, Min DS, Ahn BH, Lee YH, Suh JW, Kim JW, Iwashita S, Omori A, Ichinose S, Numata O, Seo JK, Oh YS, Suh PG (May 2002). "Interaction of elongation factor-1alpha and pleckstrin homology domain of phospholipase C-gamma 1 with activating its activity". The Journal of Biological Chemistry. 277 (22): 19697–702. doi:10.1074/jbc.M111206200. PMID 11886851.
Further reading
- Madsen HO, Poulsen K, Dahl O, Clark BF, Hjorth JP (Mar 1990). "Retropseudogenes constitute the major part of the human elongation factor 1 alpha gene family". Nucleic Acids Research. 18 (6): 1513–6. doi:10.1093/nar/18.6.1513. PMC 330519. PMID 2183196.
- Bec G, Kerjan P, Zha XD, Waller JP (Dec 1989). "Valyl-tRNA synthetase from rabbit liver. I. Purification as a heterotypic complex in association with elongation factor 1". The Journal of Biological Chemistry. 264 (35): 21131–7. doi:10.1016/S0021-9258(19)30056-0. PMID 2556394.
- Uetsuki T, Naito A, Nagata S, Kaziro Y (Apr 1989). "Isolation and characterization of the human chromosomal gene for polypeptide chain elongation factor-1 alpha". The Journal of Biological Chemistry. 264 (10): 5791–8. doi:10.1016/S0021-9258(18)83619-5. PMID 2564392.
- Whiteheart SW, Shenbagamurthi P, Chen L, Cotter RJ, Hart GW (Aug 1989). "Murine elongation factor 1 alpha (EF-1 alpha) is posttranslationally modified by novel amide-linked ethanolamine-phosphoglycerol moieties. Addition of ethanolamine-phosphoglycerol to specific glutamic acid residues on EF-1 alpha". The Journal of Biological Chemistry. 264 (24): 14334–41. doi:10.1016/S0021-9258(18)71682-7. PMID 2569467.
- Wolfson AD, Orlovsky AF, Gladilin KL (Oct 1988). "Mammalian valyl-tRNA synthetase forms a complex with the first elongation factor". FEBS Letters. 238 (2): 262–4. doi:10.1016/0014-5793(88)80492-7. PMID 3169261. S2CID 45934458.
- Ann DK, Wu MM, Huang T, Carlson DM, Wu R (Mar 1988). "Retinol-regulated gene expression in human tracheobronchial epithelial cells. Enhanced expression of elongation factor EF-1 alpha". The Journal of Biological Chemistry. 263 (8): 3546–9. doi:10.1016/S0021-9258(18)68958-6. PMID 3346208.
- Brands JH, Maassen JA, van Hemert FJ, Amons R, Möller W (Feb 1986). "The primary structure of the alpha subunit of human elongation factor 1. Structural aspects of guanine-nucleotide-binding sites". European Journal of Biochemistry. 155 (1): 167–71. doi:10.1111/j.1432-1033.1986.tb09472.x. PMID 3512269.
- Opdenakker G, Cabeza-Arvelaiz Y, Fiten P, Dijkmans R, Van Damme J, Volckaert G, Billiau A, Van Elsen A, Van der Schueren B, Van den Berghe H (Apr 1987). "Human elongation factor 1 alpha: a polymorphic and conserved multigene family with multiple chromosomal localizations". Human Genetics. 75 (4): 339–44. doi:10.1007/BF00284104. PMID 3570288. S2CID 19189698.
- Rao TR, Slobin LI (Mar 1986). "Structure of the amino-terminal end of mammalian elongation factor Tu". Nucleic Acids Research. 14 (5): 2409. doi:10.1093/nar/14.5.2409. PMC 339668. PMID 3960725.
- Carvalho MD, Carvalho JF, Merrick WC (Nov 1984). "Biological characterization of various forms of elongation factor 1 from rabbit reticulocytes". Archives of Biochemistry and Biophysics. 234 (2): 603–11. doi:10.1016/0003-9861(84)90310-2. PMID 6568109.
- Shen R, Su ZZ, Olsson CA, Fisher PB (Jul 1995). "Identification of the human prostatic carcinoma oncogene PTI-1 by rapid expression cloning and differential RNA display". Proceedings of the National Academy of Sciences of the United States of America. 92 (15): 6778–82. Bibcode:1995PNAS...92.6778S. doi:10.1073/pnas.92.15.6778. PMC 41412. PMID 7542776.
- Reed VS, Wastney ME, Yang DC (Dec 1994). "Mechanisms of the transfer of aminoacyl-tRNA from aminoacyl-tRNA synthetase to the elongation factor 1 alpha". The Journal of Biological Chemistry. 269 (52): 32932–6. doi:10.1016/S0021-9258(20)30080-6. PMID 7806521.
- Kielbassa K, Müller HJ, Meyer HE, Marks F, Gschwendt M (Mar 1995). "Protein kinase C delta-specific phosphorylation of the elongation factor eEF-alpha and an eEF-1 alpha peptide at threonine 431". The Journal of Biological Chemistry. 270 (11): 6156–62. doi:10.1074/jbc.270.11.6156. PMID 7890750.
- Maruyama K, Sugano S (Jan 1994). "Oligo-capping: a simple method to replace the cap structure of eukaryotic mRNAs with oligoribonucleotides". Gene. 138 (1–2): 171–4. doi:10.1016/0378-1119(94)90802-8. PMID 8125298.
- Wu-Baer F, Lane WS, Gaynor RB (Feb 1996). "Identification of a group of cellular cofactors that stimulate the binding of RNA polymerase II and TRP-185 to human immunodeficiency virus 1 TAR RNA". The Journal of Biological Chemistry. 271 (8): 4201–8. doi:10.1074/jbc.271.8.4201. PMID 8626763.
- Sanders J, Brandsma M, Janssen GM, Dijk J, Möller W (May 1996). "Immunofluorescence studies of human fibroblasts demonstrate the presence of the complex of elongation factor-1 beta gamma delta in the endoplasmic reticulum". Journal of Cell Science. 109 (5): 1113–7. doi:10.1242/jcs.109.5.1113. hdl:1854/LU-01GJAXGWEXX2G4TPB1AKB720BQ. PMID 8743958.
- Welle S, Thornton C, Bhatt K, Krym M (Sep 1997). "Expression of elongation factor-1 alpha and S1 in young and old human skeletal muscle". The Journals of Gerontology. Series A, Biological Sciences and Medical Sciences. 52 (5): B235-9. doi:10.1093/gerona/52A.5.B235. PMID 9310071.
- Suzuki Y, Yoshitomo-Nakagawa K, Maruyama K, Suyama A, Sugano S (Oct 1997). "Construction and characterization of a full length-enriched and a 5'-end-enriched cDNA library". Gene. 200 (1–2): 149–56. doi:10.1016/S0378-1119(97)00411-3. PMID 9373149.
- Sheu GT, Traugh JA (Dec 1997). "Recombinant subunits of mammalian elongation factor 1 expressed in Escherichia coli. Subunit interactions, elongation activity, and phosphorylation by protein kinase CKII". The Journal of Biological Chemistry. 272 (52): 33290–7. doi:10.1074/jbc.272.52.33290. PMID 9407120.