Protease-activated receptor
Protease-activated receptors (PAR) are a subfamily of related G protein-coupled receptors that are activated by cleavage of part of their extracellular domain. They are highly expressed in platelets, and also on endothelial cells, fibroblasts, immune cells, myocytes, neurons, and tissues that line the gastrointestinal tract.[1]
| coagulation factor II thrombin receptor | |||||||
|---|---|---|---|---|---|---|---|
| Identifiers | |||||||
| Symbol | F2R | ||||||
| Alt. symbols | TR; HTR; CF2R; PAR1; PAR-1 | ||||||
| NCBI gene | 2149 | ||||||
| HGNC | 3537 | ||||||
| OMIM | 187930 | ||||||
| PDB | 3BEF | ||||||
| RefSeq | NM_001992 | ||||||
| UniProt | P25116 | ||||||
| Other data | |||||||
| Locus | Chr. 5 q13.3 | ||||||
| |||||||
| F2R like trypsin receptor 1 | |||||||
|---|---|---|---|---|---|---|---|
| Identifiers | |||||||
| Symbol | F2RL1 | ||||||
| Alt. symbols | PAR2, GPR11 | ||||||
| NCBI gene | 2150 | ||||||
| HGNC | 3538 | ||||||
| OMIM | 600933 | ||||||
| PDB | 5NDD | ||||||
| RefSeq | NM_005242 | ||||||
| UniProt | P55085 | ||||||
| Other data | |||||||
| Locus | Chr. 5 q13.3 | ||||||
| |||||||
| coagulation factor II thrombin receptor like 2 | |||||||
|---|---|---|---|---|---|---|---|
| Identifiers | |||||||
| Symbol | F2RL2 | ||||||
| Alt. symbols | PAR3; PAR-3 | ||||||
| NCBI gene | 2151 | ||||||
| HGNC | 3539 | ||||||
| OMIM | 601919 | ||||||
| PDB | 2PUX | ||||||
| RefSeq | NM_004101 | ||||||
| UniProt | O00254 | ||||||
| Other data | |||||||
| Locus | Chr. 5 q13.3 | ||||||
| |||||||
| F2R like thrombin or trypsin receptor 3 | |||||||
|---|---|---|---|---|---|---|---|
| Identifiers | |||||||
| Symbol | F2RL3 | ||||||
| Alt. symbols | PAR4 | ||||||
| NCBI gene | 9002 | ||||||
| HGNC | 3540 | ||||||
| OMIM | 602779 | ||||||
| PDB | 2PV9 | ||||||
| RefSeq | NM_003950 | ||||||
| UniProt | Q96RI0 | ||||||
| Other data | |||||||
| Locus | Chr. 19 p13.11 | ||||||
| |||||||
Protease-activated receptors PAR are not to be mistaken with PAR proteins, a group of regulators of cellular polarity[2] named after their associated partitioning phenotype.[3]
Classification
There are four mammalian members of the protease-activated receptor (PAR) family: PAR1 - encoded by the gene F2R, PAR2 - F2RL1, PAR3 - F2RL2 and PAR4 - F2RL3, all these genes have their locus on chromosome 5 except of PAR4, which is on chromosome 19. The protease specificities, expression patterns, and functions of each PAR vary across a range of tissues and cell types. They are also members of the seven-transmembrane G-protein-coupled receptor superfamily, and are expressed throughout the body.[4]
History
When scientists were researching the process of blood clotting in the late 1980s, they made the discovery of protease-activated receptors (PARs). A novel protein that was activated by thrombin, a crucial part of the clotting cascade, was discovered by a research team at the University of California, San Francisco in 1991. The team was directed by Shaun Coughlin. This protein, which was eventually given the designation protease-activated receptor 1 (PAR1), was the first to be recognized as a member of the PAR family.
A second thrombin-activated protein, later known as PAR3, was identified in 1994. Later research revealed that PAR3 works as a cofactor for PAR4 but lacks a useful intracellular domain. A third member of the PAR family, known as PAR2, was discovered as a protein triggered by in 1996.
Activation
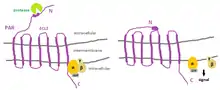
Protease activated receptors are integral membrane proteins that are coupled to G-proteins and are activated by proteolytic cleavage of the amino terminal sequence that exposes a new N-terminal sequence functions as a tethered ligand, which bind a conserved region on extracellular loop 2 (ECL2). Such binding causes activation of intracellular pathways and the specific change in conformation of the PAR. The most effective activator of PAR4 is thrombin. However, trypsin and factor Xa activate PAR4 by cleaving the receptor at various places in its N-terminal domain.[4] Four types of PAR receptors have been identified by molecular cloning, and classified according to the main enzyme that is able to activate it. It has been determined that a large group of proteases cleave and activate PARs receptors, including various endogenous proteases from: a) the coagulation cascade, b) inflammatory cells, and c) the digestive tract. On the other hand, PARs can be specifically cleaved and irreversibly activated even by exogenous proteases originated from insects, bacteria or plants and fungi.[4] The wide distribution of PARs in a variety of cells supports the idea that they are involved in many process related with the gastrointestinal physiology and cardiovascular physiology.[5] Although the proteolysis is the main mechanism for PAR activation, it is well known that a synthetic peptide (SLIGKV) that mimics the new N-terminal sequence produced after the cleavage, activates PAR-2 receptors without its proteolytic processing. In this sense, here we report that TFF3 isolated from human breast milk activates PAR-2 receptors of intestinal epithelial cells HT-29. These findings suggest that TFF3 activates intestinal epithelial cells through G-protein-coupled PAR-2, and could actively participate in the immune system of breastfed babies inducing the production of peptides related to innate defense, such as defensins and cytokines.[5]
PARs are activated by the action of serine proteases such as thrombin (acts on PARs 1, 3 and 4) and trypsin (PAR 2).[6] These enzymes cleave the N-terminus of the receptor, which in turn acts as a tethered ligand. In the cleaved state, part of the receptor itself acts as the agonist, causing a physiological response.
Most of the PAR family act through the actions of G-proteins i (cAMP inhibitory), 12/13 (Rho and Ras activation) and q (calcium signalling) to cause cellular actions.
Function
PARs play a role in a multitude of physiological processes such as hemostasis, thrombosis, inflammation, and pain sensation. The cellular effects of thrombin are mediated by protease-activated receptors (PARs). Endothelial PARs participate in the regulation of vascular tone and permeability while in vascular smooth muscle they mediate contraction, proliferation, and hypertrophy. In endothelial cells PARs play a key role in promotion vascular barrier function as they provide a positive signals for endothelial adhesion molecules (vascular cell adhesion molecule-1 (VCAM-1), intercellular adhesion molecule-1(ICAM-1), and E-selectin).[7] PARs contribute to the pro-inflammatory response. For example PAR4 induces leukocyte migration and PAR2 helps macrophages to produce cytokines such as interleukin-8 (IL-8). Recent research has also implicated these novel receptors in muscle growth and bone cell differentiation and proliferation.[4]
References
- Macfarlane SR, Seatter MJ, Kanke T, Hunter GD, Plevin R (June 2001). "Proteinase-activated receptors" (abstract). Pharmacological Reviews. 53 (2): 245–282. PMID 11356985.
- Munro EM (February 2006). "PAR proteins and the cytoskeleton: a marriage of equals". Current Opinion in Cell Biology. 18 (1): 86–94. doi:10.1016/j.ceb.2005.12.007. PMID 16364625.
- Goldstein B, Macara IG (November 2007). "The PAR proteins: fundamental players in animal cell polarization". Developmental Cell. 13 (5): 609–622. doi:10.1016/j.devcel.2007.10.007. PMC 2964935. PMID 17981131.
- Heuberger DM, Schuepbach RA (December 2019). "Protease-activated receptors (PARs): mechanisms of action and potential therapeutic modulators in PAR-driven inflammatory diseases". Thrombosis Journal. 17 (1): 4. doi:10.1186/s12959-019-0194-8. PMC 6440139. PMID 30976204.
- Barrera GJ, Tortolero GS (2016). "Trefoil factor 3 (TFF3) from human breast milk activates PAR-2 receptors, of the intestinal epithelial cells HT-29, regulating cytokines and defensins". Bratislavske Lekarske Listy. 117 (6): 332–339. doi:10.4149/bll_2016_066. PMID 27546365.
- Pawar NR, Buzza MS, Antalis TM (January 2019). "Membrane-Anchored Serine Proteases and Protease-Activated Receptor-2-Mediated Signaling: Co-Conspirators in Cancer Progression". Cancer Research. 79 (2): 301–310. doi:10.1158/0008-5472.CAN-18-1745. PMC 6335149. PMID 30610085.
- Bae JS, Rezaie AR (March 2009). "Thrombin inhibits nuclear factor kappaB and RhoA pathways in cytokine-stimulated vascular endothelial cells when EPCR is occupied by protein C". Thrombosis and Haemostasis. 101 (3): 513–520. doi:10.1160/th08-09-0568. PMC 2688729. PMID 19277413.
Further reading
- Macfarlane SR, Seatter MJ, Kanke T, Hunter GD, Plevin R (June 2001). "Proteinase-activated receptors". Pharmacological Reviews. 53 (2): 245–282. PMID 11356985.
- Hollenberg MD, Compton SJ (June 2002). "International Union of Pharmacology. XXVIII. Proteinase-activated receptors". Pharmacological Reviews. 54 (2): 203–217. doi:10.1124/pr.54.2.203. PMID 12037136. S2CID 3188040.
- Coughlin SR (August 2005). "Protease-activated receptors in hemostasis, thrombosis and vascular biology". Journal of Thrombosis and Haemostasis. 3 (8): 1800–1814. doi:10.1111/j.1538-7836.2005.01377.x. PMID 16102047. S2CID 38430621.
- Adams MN, Ramachandran R, Yau MK, Suen JY, Fairlie DP, Hollenberg MD, Hooper JD (June 2011). "Structure, function and pathophysiology of protease activated receptors". Pharmacology & Therapeutics. 130 (3): 248–282. doi:10.1016/j.pharmthera.2011.01.003. PMID 21277892.
External links
- "Protease-Activated Receptors". IUPHAR Database of Receptors and Ion Channels. International Union of Basic and Clinical Pharmacology.
- Proteinase-Activated+Receptors at the U.S. National Library of Medicine Medical Subject Headings (MeSH)
- Protease activated receptor peptides