Cannabinoid receptor 1
Cannabinoid receptor 1 (CB1), is a G protein-coupled cannabinoid receptor that in humans is encoded by the CNR1 gene.[5] The human CB1 receptor is expressed in the peripheral nervous system and central nervous system.[5] It is activated by endocannabinoids, a group of retrograde neurotransmitters that include anandamide and 2-arachidonoylglycerol (2-AG); plant phytocannabinoids, such as docosatetraenoylethanolamide found in wild daga, the compound THC which is an active constituent of the psychoactive drug cannabis; and synthetic analogs of THC. CB1 is antagonized by the phytocannabinoid tetrahydrocannabivarin (THCV).[6][7]
_PDB_5XRA.png.webp)


The primary endogenous agonist of the human CB1 receptor is anandamide.[5]
Structure
The CB1 receptor shares the structure characteristic of all G-protein-coupled receptors, possessing seven transmembrane domains connected by three extracellular and three intracellular loops, an extracellular N-terminal tail, and an intracellular C-terminal tail.[8][9] The receptor may exist as a homodimer or form heterodimers or other GPCR oligomers with different classes of G-protein-coupled receptors. Observed heterodimers include A2A–CB1, CB1–D2, OX1–CB1, μOR–CB1, while many more may only be stable enough to exist in vivo.[10][11] The CB1 receptor possesses an allosteric modulatory binding site.[12][13]
Mechanism
The CB1 receptor is a pre-synaptic heteroreceptor that modulates neurotransmitter release when activated in a dose-dependent, stereoselective and pertussis toxin-sensitive manner.[14] The CB1 receptor is activated by cannabinoids, generated naturally inside the body (endocannabinoids) or exogenously, normally through cannabis or a related synthetic compound.
Research suggests that the majority of CB1 receptors are coupled through Gi/o proteins. Upon activation, CB1 receptor exhibits its effects mainly through activation of Gi, which decreases intracellular cAMP concentration by inhibiting its production enzyme, adenylate cyclase, and increases mitogen-activated protein kinase (MAP kinase) concentration. Alternatively, in some rare cases CB1 receptor activation may be coupled to Gs proteins, which stimulate adenylate cyclase.[11] cAMP is known to serve as a second messenger coupled to a variety of ion channels, including the positively influenced inwardly rectifying potassium channels (=Kir or IRK),[15] and calcium channels, which are activated by cAMP-dependent interaction with such molecules as protein kinase A (PKA), protein kinase C (PKC), Raf-1, ERK, JNK, p38, c-fos, c-jun, and others.[16]
In terms of function, the inhibition of intracellular cAMP expression shortens the duration of pre-synaptic action potentials by prolonging the rectifying potassium A-type currents, which is normally inactivated upon phosphorylation by PKA. This inhibition grows more pronounced when considered with the effect of activated CB1 receptors to limit calcium entry into the cell, which does not occur through cAMP but by a direct G-protein-mediated inhibition. As presynaptic calcium entry is a requirement for vesicle release, this function will decrease the transmitter that enters the synapse upon release.[17] The relative contribution of each of these two inhibitory mechanisms depends on the variance of ion channel expression by cell type.
The CB1 receptor can also be allosterically modulated by synthetic ligands[18] in a positive[19] and negative[20] manner. In vivo exposure to THC impairs long-term potentiation and leads to a reduction of phosphorylated CREB.[21]
The signaling properties of activated CB1 are furthermore modified by the presence of SGIP1, that hinders receptor internalization and decreases ERK1/2 signalling while augmenting the interaction with GRK3, β-arrestin-2.[22][23]
In summary, CB1 receptor activity has been found to be coupled to certain ion channels, in the following manner:[11]
- Positively to inwardly rectifying and A-type outward potassium channels.
- Negatively to D-type outward potassium channels
- Negatively to N-type and P/Q-type calcium channels.
Expression
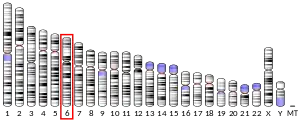
The CB1 receptor is encoded by the gene CNR1,[14] located on human chromosome 6.[17] Two transcript variants encoding different isoforms have been described for this gene.[14] CNR1 orthologs[24] have been identified in most mammals.
The CB1 receptor is expressed pre-synaptically at both glutaminergic and GABAergic interneurons and, in effect, acts as a neuromodulator to inhibit release of glutamate and GABA.[17] Repeated administration of receptor agonists may result in receptor internalization and/or a reduction in receptor protein signalling.[11]
The inverse agonist MK-9470 makes it possible to produce in vivo images of the distribution of CB1 receptors in the human brain with positron emission tomography.[25]
Brain

CB1 receptors are expressed most densely in the central nervous system and are largely responsible for mediating the effects of cannabinoid binding in the brain. Endocannabinoids released by a depolarized neuron bind to CB1 receptors on pre-synaptic glutamatergic and GABAergic neurons, resulting in a respective decrease in either glutamate or GABA release. Limiting glutamate release causes reduced excitation, while limiting GABA release suppresses inhibition, a common form of short-term plasticity in which the depolarization of a single neuron induces a reduction in GABA-mediated inhibition, in effect exciting the postsynaptic cell.[17]
Varying levels of CB1 expression can be detected in the olfactory bulb, cortical regions (neocortex, pyriform cortex, hippocampus, and amygdala), several parts of basal ganglia, thalamic and hypothalamic nuclei, and other subcortical regions (e.g., the septal region), cerebellar cortex, and brainstem nuclei (e.g., the periaqueductal gray).[16]
Hippocampal formation
CB1 mRNA transcripts are abundant in GABAergic interneurons of the hippocampus, indirectly reflecting the expression of these receptors and elucidating the established effect of cannabinoids on memory. These receptors are densely located in cornu ammonis pyramidal cells, which are known to release glutamate. Cannabinoids suppress the induction of LTP and LTD in the hippocampus by inhibiting these glutamatergic neurons. By reducing the concentration of glutamate released below the threshold necessary to depolarize the postsynaptic receptor NMDA,[17] a receptor known to be directly related to the induction of LTP and LTD, cannabinoids are a crucial factor in the selectivity of memory. These receptors are highly expressed by GABAergic interneurons as well as glutamatergic principal neurons. However, a higher density is found within GABAergic cells.[26] This means that, although synaptic strength/frequency, and thus potential to induce LTP, is lowered, net hippocampal activity is raised. In addition, CB1 receptors in the hippocampus indirectly inhibit the release of acetylcholine. This serves as the modulatory axis opposing GABA, decreasing neurotransmitter release. Cannabinoids also likely play an important role in the development of memory through their neonatal promotion of myelin formation, and thus the individual segregation of axons.
Basal ganglia
CB1 receptors are expressed throughout the basal ganglia and have well-established effects on movement in rodents. As in the hippocampus, these receptors inhibit the release of glutamate or GABA transmitter, resulting in decreased excitation or reduced inhibition based on the cell they are expressed in. Consistent with the variable expression of both excitatory glutamate and inhibitory GABA interneurons in both the basal ganglia's direct and indirect motor loops, synthetic cannabinoids are known to influence this system in a dose-dependent triphasic pattern. Decreased locomotor activity is seen at both higher and lower concentrations of applied cannabinoids, whereas an enhancement of movement may occur upon moderate dosages.[17] However, these dose-dependent effects have been studied predominately in rodents, and the physiological basis for this triphasic pattern warrants future research in humans. Effects may vary based on the site of cannabinoid application, input from higher cortical centers, and whether drug application is unilateral or bilateral.
Cerebellum and neocortex
The role of the CB1 receptor in the regulation of motor movements is complicated by the additional expression of this receptor in the cerebellum and neocortex, two regions associated with the coordination and initiation of movement. Research suggests that anandamide is synthesized by Purkinje cells and acts on presynaptic receptors to inhibit glutamate release from granule cells or GABA release from the terminals of basket cells. In the neocortex, these receptors are concentrated on local interneurons in cerebral layers II-III and V-VI.[17] Compared to rat brains, humans express more CB1 receptors in the cerebral cortex and amygdala and less in the cerebellum, which may help explain why motor function seems to be more compromised in rats than humans upon cannabinoid application.[26]
Spine
Many of the documented analgesic effects of cannabinoids are based on the interaction of these compounds with CB1 receptors on spinal cord interneurons in the superficial levels of the dorsal horn, known for its role in nociceptive processing. In particular, the CB1 is heavily expressed in layers 1 and 2 of the spinal cord dorsal horn and in lamina 10 by the central canal. Dorsal root ganglion also express these receptors, which target a variety of peripheral terminals involved in nociception. Signals on this track are also transmitted to the periaqueductal gray (PAG) of the midbrain. Endogenous cannabinoids are believed to exhibit an analgesic effect on these receptors by limiting both GABA and glutamate of PAG cells that relate to nociceptive input processing, a hypothesis consistent with the finding that anandamide release in the PAG is increased in response to pain-triggering stimuli.[17]
Other
CB1 is expressed on several types of cells in pituitary gland, thyroid gland, and possibly in the adrenal gland.[16] CB1 is also expressed in several cells relating to metabolism, such as fat cells, muscle cells, liver cells (and also in the endothelial cells, Kupffer cells and stellate cells of the liver), and in the digestive tract.[16] It is also expressed in the lungs and the kidney.
CB1 is present on Leydig cells and human sperms. In females, it is present in the ovaries, oviducts myometrium, decidua, and placenta. It has also been implicated in the proper development of the embryo.[16]
CB1 is also expressed in the retina. In the retina, they are expressed in the photoreceptors, inner plexiform, outer plexiform, bipolar cells, ganglion cells, and retinal pigment epithelium cells.[27] In the visual system, cannabinoids agonist induce a dose dependent modulation of calcium, chloride and potassium channels. This alters vertical transmission between photoreceptor, bipolar and ganglion cells. Altering vertical transmission in turn results in the way vision is perceived.[28]
Use of antagonists
Selective CB1 agonists may be used to isolate the effects of the receptor from the CB1 receptor, as most cannabinoids and endocannabinoids bind to both receptor types.[17] CB1 selective antagonists are used for weight reduction and smoking cessation (see Rimonabant). A substantial number of antagonists of the CB1 receptor have been discovered and characterized. TM38837 has been developed as a CB1 receptor antagonist that is restricted to targeting only peripheral CB1 receptors.
Ligands
Agonists
Selective
Unspecified efficacy
Endogenous
Endogenous
Synthetic
Allosteric agonist
- GAT228[31]
Antagonists
- Cannabigerol
- Ibipinabant
- Otenabant
- Tetrahydrocannabivarin
- Virodhamine (Endogenous CB1 antagonist and CB2 agonist)
Inverse agonists
Allosteric modulators
- Lipoxin A4 – endogenous, PAM
- ZCZ-011 – PAM
- Pregnenolone – endogenous, NAM
- Cannabidiol – NAM[26]
- Fenofibrate – NAM
- GAT100 – NAM
- PSNCBAM-1 – NAM
- RVD-Hpα – NAM
Binding affinities
| CB1 affinity (Ki) | Efficacy towards CB1 | CB2 affinity (Ki) | Efficacy towards CB2 | Type | References | |
|---|---|---|---|---|---|---|
| Anandamide | 78 nM | Partial agonist | 370 nM | Partial agonist | Endogenous | |
| N-Arachidonoyl dopamine | 250 nM | Agonist | 12000 nM | ? | Endogenous | [32] |
| 2-Arachidonoylglycerol | 58.3 nM | Full agonist | 145 nM | Full agonist | Endogenous | [32] |
| 2-Arachidonyl glyceryl ether | 21 nM | Full agonist | 480 nM | Full agonist | Endogenous | |
| Tetrahydrocannabinol | 10 nM | Partial agonist | 24 nM | Partial agonist | Phytogenic | [33] |
| EGCG | 33600 nM | Agonist | 50000+ nM | ? | Phytogenic | |
| AM-1221 | 52.3 nM | Agonist | 0.28 nM | Agonist | Synthetic | [34] |
| AM-1235 | 1.5 nM | Agonist | 20.4 nM | Agonist | Synthetic | [35] |
| AM-2232 | 0.28 nM | Agonist | 1.48 nM | Agonist | Synthetic | [35] |
| UR-144 | 150 nM | Full agonist | 1.8 nM | Full agonist | Synthetic | [36] |
| JWH-007 | 9.0 nM | Agonist | 2.94 nM | Agonist | Synthetic | [37] |
| JWH-015 | 383 nM | Agonist | 13.8 nM | Agonist | Synthetic | [37] |
| JWH-018 | 9.00 ± 5.00 nM | Full agonist | 2.94 ± 2.65 nM | Full agonist | Synthetic | [38] |
Evolution
The CNR1 gene is used in animals as a nuclear DNA phylogenetic marker.[24] This intronless gene has first been used to explore the phylogeny of the major groups of mammals,[39] and contributed to reveal that placental orders are distributed into five major clades: Xenarthra, Afrotheria, Laurasiatheria, Euarchonta, and Glires. CNR1 has also proven useful at lower taxonomic levels, such as rodents,[40][41] and for the identification of dermopterans as the closest primate relatives.[42]
See also
References
- GRCh38: Ensembl release 89: ENSG00000118432 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000044288 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Abood M, Barth F, Bonner TI, Cabral G, Casellas P, Cravatt BF, Devane WA, Elphick MR, Felder CC, Herkenham M, Howlett AC, Kunos G, Mackie K, Mechoulam R, Pertwee RG (22 August 2018). "CB1 Receptor". IUPHAR/BPS Guide to Pharmacology. International Union of Basic and Clinical Pharmacology. Retrieved 9 November 2018.
- Thomas A, Stevenson LA, Wease KN, Price MR, Baillie G, Ross RA, Pertwee RG (December 2005). "Evidence that the plant cannabinoid Delta9-tetrahydrocannabivarin is a cannabinoid CB1 and CB2 receptor antagonist". British Journal of Pharmacology. 146 (7): 917–926. doi:10.1038/sj.bjp.0706414. PMC 1751228. PMID 16205722.
- Pertwee RG, Thomas A, Stevenson LA, Ross RA, Varvel SA, Lichtman AH, et al. (March 2007). "The psychoactive plant cannabinoid, Delta9-tetrahydrocannabinol, is antagonized by Delta8- and Delta9-tetrahydrocannabivarin in mice in vivo". British Journal of Pharmacology. 150 (5): 586–594. doi:10.1038/sj.bjp.0707124. PMC 2189766. PMID 17245367.
- Shao Z, Yin J, Chapman K, Grzemska M, Clark L, Wang J, Rosenbaum DM (December 2016). "High-resolution crystal structure of the human CB1 cannabinoid receptor". Nature. 540 (7634): 602–606. Bibcode:2016Natur.540..602S. doi:10.1038/nature20613. PMC 5433929. PMID 27851727.
- Hua T, Vemuri K, Pu M, Qu L, Han GW, Wu Y, et al. (October 2016). "Crystal Structure of the Human Cannabinoid Receptor CB1". Cell. 167 (3): 750–762.e14. doi:10.1016/j.cell.2016.10.004. PMC 5322940. PMID 27768894.
- Hojo M, Sudo Y, Ando Y, Minami K, Takada M, Matsubara T, et al. (November 2008). "mu-Opioid receptor forms a functional heterodimer with cannabinoid CB1 receptor: electrophysiological and FRET assay analysis". Journal of Pharmacological Sciences. 108 (3): 308–319. doi:10.1254/jphs.08244FP. PMID 19008645.
- Pertwee RG (April 2006). "The pharmacology of cannabinoid receptors and their ligands: an overview". International Journal of Obesity. 30 Suppl 1: S13–S18. doi:10.1038/sj.ijo.0803272. PMID 16570099.
- Nickols HH, Conn PJ (January 2014). "Development of allosteric modulators of GPCRs for treatment of CNS disorders". Neurobiology of Disease. 61: 55–71. doi:10.1016/j.nbd.2013.09.013. PMC 3875303. PMID 24076101.
- Nguyen T, Li JX, Thomas BF, Wiley JL, Kenakin TP, Zhang Y (May 2017). "Allosteric Modulation: An Alternate Approach Targeting the Cannabinoid CB1 Receptor". Medicinal Research Reviews. 37 (3): 441–474. doi:10.1002/med.21418. PMC 5397374. PMID 27879006.
- "Entrez Gene: CNR1 cannabinoid receptor 1 (brain)".
- Demuth DG, Molleman A (January 2006). "Cannabinoid signalling". Life Sciences. 78 (6): 549–563. doi:10.1016/j.lfs.2005.05.055. PMID 16109430.
- Pagotto U, Marsicano G, Cota D, Lutz B, Pasquali R (February 2006). "The emerging role of the endocannabinoid system in endocrine regulation and energy balance". Endocrine Reviews. 27 (1): 73–100. doi:10.1210/er.2005-0009. PMID 16306385.
- Elphick MR, Egertová M (March 2001). "The neurobiology and evolution of cannabinoid signalling". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 356 (1407): 381–408. doi:10.1098/rstb.2000.0787. PMC 1088434. PMID 11316486.
- Price MR, Baillie GL, Thomas A, Stevenson LA, Easson M, Goodwin R, et al. (November 2005). "Allosteric modulation of the cannabinoid CB1 receptor". Molecular Pharmacology. 68 (5): 1484–1495. doi:10.1124/mol.105.016162. PMID 16113085. S2CID 17648541.
- Navarro HA, Howard JL, Pollard GT, Carroll FI (April 2009). "Positive allosteric modulation of the human cannabinoid (CB) receptor by RTI-371, a selective inhibitor of the dopamine transporter". British Journal of Pharmacology. 156 (7): 1178–1184. doi:10.1111/j.1476-5381.2009.00124.x. PMC 2697692. PMID 19226282.
- Horswill JG, Bali U, Shaaban S, Keily JF, Jeevaratnam P, Babbs AJ, et al. (November 2007). "PSNCBAM-1, a novel allosteric antagonist at cannabinoid CB1 receptors with hypophagic effects in rats". British Journal of Pharmacology. 152 (5): 805–814. doi:10.1038/sj.bjp.0707347. PMC 2190018. PMID 17592509.
- Fan N, Yang H, Zhang J, Chen C (February 2010). "Reduced expression of glutamate receptors and phosphorylation of CREB are responsible for in vivo Delta9-THC exposure-impaired hippocampal synaptic plasticity". Journal of Neurochemistry. 112 (3): 691–702. doi:10.1111/j.1471-4159.2009.06489.x. PMC 2809144. PMID 19912468.
- Gazdarica M, Noda J, Durydivka O, Novosadova V, Mackie K, Pin JP, et al. (March 2022). "SGIP1 modulates kinetics and interactions of the cannabinoid receptor 1 and G protein-coupled receptor kinase 3 signalosome". Journal of Neurochemistry. 160 (6): 625–642. doi:10.1111/jnc.15569. PMC 9306533. PMID 34970999. S2CID 245593712.
- Hájková A, Techlovská Š, Dvořáková M, Chambers JN, Kumpošt J, Hubálková P, et al. (August 2016). "SGIP1 alters internalization and modulates signaling of activated cannabinoid receptor 1 in a biased manner". Neuropharmacology. 107: 201–214. doi:10.1016/j.neuropharm.2016.03.008. PMID 26970018. S2CID 15003497.
- "OrthoMaM phylogenetic marker: CNR1 coding sequence". Archived from the original on 22 December 2015. Retrieved 23 November 2009.
- Burns HD, Van Laere K, Sanabria-Bohórquez S, Hamill TG, Bormans G, Eng WS, et al. (June 2007). "[18F]MK-9470, a positron emission tomography (PET) tracer for in vivo human PET brain imaging of the cannabinoid-1 receptor". Proceedings of the National Academy of Sciences of the United States of America. 104 (23): 9800–9805. Bibcode:2007PNAS..104.9800B. doi:10.1073/pnas.0703472104. PMC 1877985. PMID 17535893.
- Pertwee RG (January 2008). "The diverse CB1 and CB2 receptor pharmacology of three plant cannabinoids: delta9-tetrahydrocannabinol, cannabidiol and delta9-tetrahydrocannabivarin". British Journal of Pharmacology. 153 (2): 199–215. doi:10.1038/sj.bjp.0707442. PMC 2219532. PMID 17828291.
- Porcella A, Maxia C, Gessa GL, Pani L (March 2000). "The human eye expresses high levels of CB1 cannabinoid receptor mRNA and protein". The European Journal of Neuroscience. 12 (3): 1123–1127. doi:10.1046/j.1460-9568.2000.01027.x. PMID 10762343. S2CID 34849187.
- Hoon M, Okawa H, Della Santina L, Wong RO (September 2014). "Functional architecture of the retina: development and disease". Progress in Retinal and Eye Research. 42: 44–84. doi:10.1016/j.preteyeres.2014.06.003. PMC 4134977. PMID 24984227.
- Lopez-Rodriguez AB, Siopi E, Finn DP, Marchand-Leroux C, Garcia-Segura LM, Jafarian-Tehrani M, Viveros MP (January 2015). "CB1 and CB2 cannabinoid receptor antagonists prevent minocycline-induced neuroprotection following traumatic brain injury in mice". Cerebral Cortex. 25 (1): 35–45. doi:10.1093/cercor/bht202. PMID 23960212.
- Leggett JD, Aspley S, Beckett SR, D'Antona AM, Kendall DA, Kendall DA (January 2004). "Oleamide is a selective endogenous agonist of rat and human CB1 cannabinoid receptors". British Journal of Pharmacology. 141 (2): 253–262. doi:10.1038/sj.bjp.0705607. PMC 1574194. PMID 14707029.
- Laprairie RB, Kulkarni PM, Deschamps JR, Kelly ME, Janero DR, Cascio MG, et al. (June 2017). "Enantiospecific Allosteric Modulation of Cannabinoid 1 Receptor". ACS Chemical Neuroscience. 8 (6): 1188–1203. doi:10.1021/acschemneuro.6b00310. PMID 28103441.
- Pertwee RG, Howlett AC, Abood ME, Alexander SP, Di Marzo V, Elphick MR, et al. (December 2010). "International Union of Basic and Clinical Pharmacology. LXXIX. Cannabinoid receptors and their ligands: beyond CB₁ and CB₂". Pharmacological Reviews. 62 (4): 588–631. doi:10.1124/pr.110.003004. PMC 2993256. PMID 21079038.
- "PDSP Database – UNC". Archived from the original on 8 November 2013. Retrieved 11 June 2013.
- WO patent 200128557, Makriyannis A, Deng H, "Cannabimimetic indole derivatives", granted 2001-06-07
- US patent 7241799, Makriyannis A, Deng H, "Cannabimimetic indole derivatives", granted 2007-07-10
- Frost JM, Dart MJ, Tietje KR, Garrison TR, Grayson GK, Daza AV, et al. (January 2010). "Indol-3-ylcycloalkyl ketones: effects of N1 substituted indole side chain variations on CB(2) cannabinoid receptor activity". Journal of Medicinal Chemistry. 53 (1): 295–315. doi:10.1021/jm901214q. PMID 19921781.
- Aung MM, Griffin G, Huffman JW, Wu M, Keel C, Yang B, et al. (August 2000). "Influence of the N-1 alkyl chain length of cannabimimetic indoles upon CB(1) and CB(2) receptor binding". Drug and Alcohol Dependence. 60 (2): 133–140. doi:10.1016/S0376-8716(99)00152-0. PMID 10940540.
- Aung MM, Griffin G, Huffman JW, Wu M, Keel C, Yang B, et al. (August 2000). "Influence of the N-1 alkyl chain length of cannabimimetic indoles upon CB(1) and CB(2) receptor binding". Drug and Alcohol Dependence. 60 (2): 133–140. doi:10.1016/s0376-8716(99)00152-0. PMID 10940540.
- Murphy WJ, Eizirik E, Johnson WE, Zhang YP, Ryder OA, O'Brien SJ (February 2001). "Molecular phylogenetics and the origins of placental mammals". Nature. 409 (6820): 614–618. Bibcode:2001Natur.409..614M. doi:10.1038/35054550. PMID 11214319. S2CID 4373847.
- Blanga-Kanfi S, Miranda H, Penn O, Pupko T, DeBry RW, Huchon D (April 2009). "Rodent phylogeny revised: analysis of six nuclear genes from all major rodent clades". BMC Evolutionary Biology. 9: 71. doi:10.1186/1471-2148-9-71. PMC 2674048. PMID 19341461.
- DeBry RW (October 2003). "Identifying conflicting signal in a multigene analysis reveals a highly resolved tree: the phylogeny of Rodentia (Mammalia)". Systematic Biology. 52 (5): 604–617. doi:10.1080/10635150390235403. PMID 14530129.
- Janecka JE, Miller W, Pringle TH, Wiens F, Zitzmann A, Helgen KM, et al. (November 2007). "Molecular and genomic data identify the closest living relative of primates". Science. 318 (5851): 792–794. Bibcode:2007Sci...318..792J. doi:10.1126/science.1147555. PMID 17975064. S2CID 12251814.
External links
- "Cannabinoid Receptors: CB1". IUPHAR Database of Receptors and Ion Channels. International Union of Basic and Clinical Pharmacology. Archived from the original on 5 March 2012.
- Cannabinoid receptor 1 (CNR1) Human Protein Atlas
This article incorporates text from the United States National Library of Medicine, which is in the public domain.