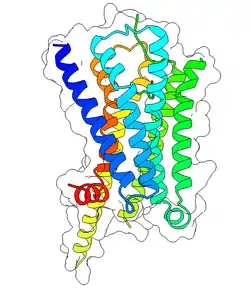
5-HT1A receptor
The serotonin 1A receptor (or 5-HT1A receptor) is a subtype of serotonin receptors, or 5-HT receptors, that binds serotonin, also known as 5-HT, a neurotransmitter. 5-HT1A is expressed in the brain, spleen, and neonatal kidney. It is a G protein-coupled receptor (GPCR), coupled to the Gi protein, and its activation in the brain mediates hyperpolarization and reduction of firing rate of the postsynaptic neuron. In humans, the serotonin 1A receptor is encoded by the HTR1A gene.[5][6]


Distribution
The 5-HT1A receptor is the most widespread of all the 5-HT receptors. In the central nervous system, 5-HT1A receptors exist in the cerebral cortex, hippocampus, septum, amygdala, and raphe nucleus in high densities, while low amounts also exist in the basal ganglia and thalamus.[7][8][9] The 5-HT1A receptors in the raphe nucleus are largely somatodendritic autoreceptors, whereas those in other areas such as the hippocampus are postsynaptic receptors.[8]
Function
Neuromodulation
5-HT1A receptor agonists are involved in neuromodulation. They decrease blood pressure and heart rate via a central mechanism, by inducing peripheral vasodilation, and by stimulating the vagus nerve.[10] These effects are the result of activation of 5-HT1A receptors within the rostral ventrolateral medulla.[10] The sympatholytic antihypertensive drug urapidil is an α1-adrenergic receptor antagonist and 5-HT1A receptor agonist, and it has been demonstrated that the latter property contributes to its overall therapeutic effects.[11][12] Vasodilation of the blood vessels in the skin via central 5-HT1A activation increases heat dissipation from the organism out into the environment, causing a decrease in body temperature.[13][14]
Activation of central 5-HT1A receptors triggers the release or inhibition of norepinephrine depending on species, presumably from the locus coeruleus, which then reduces or increases neuronal tone to the iris sphincter muscle by modulation of postsynaptic α2-adrenergic receptors within the Edinger-Westphal nucleus, resulting in pupil dilation in rodents, and pupil constriction in primates including humans.[15][16][17]
5-HT1A receptor agonists like buspirone[18] and flesinoxan[19] show efficacy in relieving anxiety[20] and depression,[21] and buspirone and tandospirone are currently approved for these indications in various parts of the world. Others such as gepirone,[22] flesinoxan,[19] flibanserin,[23] and naluzotan[24] have also been investigated, though none have been fully developed and approved yet. Some of the atypical antipsychotics like lurasidone[25] and aripiprazole[26] are also partial agonists at the 5-HT1A receptor and are sometimes used in low doses as augmentations to standard antidepressants like the selective serotonin reuptake inhibitors (SSRIs).[27] Mice lacking 5-HT1A receptors altogether (knockout) show increased anxiety but lower depressive-like behaviour.[28]
5-HT1A autoreceptor desensitization and increased 5-HT1A receptor postsynaptic activation via general increases in serotonin levels by serotonin precursor supplementation, serotonin reuptake inhibition, or inhibition of monoamine oxidase has been shown to be a major mediator in the therapeutic benefits of most mainstream antidepressant supplements and pharmaceuticals, including serotonin precursors like L-tryptophan and 5-HTP, SSRIs, serotonin-norepinephrine reuptake inhibitors (SNRIs), tricyclic antidepressants (TCAs), tetracyclic antidepressants (TeCAs), and monoamine oxidase inhibitors (MAOIs).[29] 5-HT1A receptor activation likely plays a significant role in the positive effects of serotonin releasing agents (SRAs) like MDMA (commonly known as ecstasy) as well.[30][31]
5-HT1A receptors in the dorsal raphe nucleus are co-localized with neurokinin 1 (NK1) receptors and have been shown to inhibit the release of substance P, their endogenous ligand.[32][33] In addition to being antidepressant and anxiolytic in effect, 5-HT1A receptor activation has also been demonstrated to be antiemetic[34][35] and analgesic,[36][37] and all of these properties may be mediated in part or full, depending on the property in question, by NK1 receptor inhibition. Consequently, novel NK1 receptor antagonists are now in use for the treatment of nausea and emesis, and are also being investigated for the treatment of anxiety and depression.[38]
5-HT1A receptor activation has been shown to increase dopamine release in the medial prefrontal cortex, striatum, and hippocampus, and may be useful for improving the symptoms of schizophrenia and Parkinson's disease.[39] As mentioned above, some of the atypical antipsychotics are 5-HT1A receptor partial agonists, and this property has been shown to enhance their clinical efficacy.[39][40][41] Enhancement of dopamine release in these areas may also play a major role in the antidepressant and anxiolytic effects seen upon postsynaptic activation of the 5-HT1A receptor.[42][43]
The activation of 5-HT1A receptors has been demonstrated to impair certain aspects of memory (affecting declarative and non-declarative memory functions) and learning (due to interference with memory-encoding mechanisms), by inhibiting the release of glutamate and acetylcholine in various areas of the brain.[44] 5-HT1A activation is known to improve cognitive functions associated with the prefrontal cortex, possibly via inducing prefrontal cortex dopamine and acetylcholine release.[45] Conversely, the 5-HT1A antagonist, WAY100635, alleviated learning and memory impairments induced by glutamate blockade (with dizocilpine)[46] or hippocampal cholinergic denervation (by fornix transection)[47] in primates. Furthermore, 5-HT1A receptor antagonists such as lecozotan have been shown to facilitate certain types of learning and memory in rodents, and as a result, are being developed as novel treatments for Alzheimer's disease.[48]
Other effects of 5-HT1A activation that have been observed in scientific research include:
- Decreased aggression[49][50]
- Increased sociability[31]
- Decreased impulsivity[51]
- Inhibition of drug-seeking behavior[52][53][54]
- Facilitation of sex drive and arousal[55][56]
- Inhibition of penile erection[57][58]
- Diminished food intake[59]
- Prolongation of REM sleep latency[60][61]
- Reversal of opioid-induced respiratory depression[62]
Endocrinology
5-HT1A receptor activation induces the secretion of various hormones including cortisol, corticosterone, adrenocorticotropic hormone (ACTH), oxytocin, prolactin, growth hormone, and β-endorphin.[63][64][65][66] The receptor does not affect vasopressin or renin secretion, unlike the 5-HT2 receptors.[63][64] It has been suggested that oxytocin release may contribute to the prosocial, antiaggressive, and anxiolytic properties observed upon activation of the receptor.[31] β-Endorphin secretion may contribute to antidepressant, anxiolytic, and analgesic effects.[67]
Autoreceptors
5-HT1A receptors can be located on the cell body, dendrites, axons, and both presynaptically and postsynaptically in nerve terminals or synapses. Those located on the soma and dendrites are referred to as somatodendritic, and those located presynaptically in the synapse are simply referred to as presynaptic. As a group, receptors that are sensitive to the neurotransmitter that is released by the neuron on which the receptors are located are known as autoreceptors; they typically constitute the key component of an ultra-short negative feedback loop whereby the neuron's release of neurotransmitter inhibits its further release of neurotransmitter. Stimulation of 5-HT1A autoreceptors inhibits the release of serotonin in nerve terminals. For this reason, 5-HT1A receptor agonists tend to exert a biphasic mode of action; they decrease serotonin release and postsynaptic 5-HT1A receptor activity in low doses, and further decrease serotonin release but increase postsynaptic 5-HT1A receptor activity at higher doses by directly stimulating the receptors in place of serotonin.
This autoreceptor-mediated inhibition of serotonin release has been theorized to be a major factor in the therapeutic lag that is seen with serotonergic antidepressants such as the SSRIs.[68] The autoreceptors must first desensitize before the concentration of extracellular serotonin in the synapse can become elevated appreciably.[68][69] Though the responsiveness of the autoreceptors is somewhat reduced with chronic treatment, they still remain effective at constraining large increases in extracellular serotonin concentrations.[68] For this reason, serotonin reuptake inhibitors that also have 5-HT1A receptor antagonistic or partial agonistic properties, such as vilazodone and SB-649,915, are being investigated and introduced as novel antidepressants with the potential for a faster onset of action and improved effectiveness compared to those currently available.[70]
Unlike most drugs that elevate extracellular serotonin levels like the SSRIs and MAOIs, SRAs such as fenfluramine and MDMA bypass serotonin autoreceptors such as 5-HT1A. They do this by directly acting on the release mechanisms of serotonin neurons and forcing release to occur regardless of autoreceptor-mediated inhibition.[71] As such, SRAs induce immediate and much greater increases in extracellular serotonin concentrations compared to other serotonin-elevating agents such as the SSRIs. [Note: This is questionable as the level of serotonin output from SRAs is still dose dependant and, while SRAs will initially bypass autoreceptors, the increase in serotonin they induce will then agonise autoreceptors.] In contrast to SRAs, SSRIs may decrease serotonin levels initially (especially at lower dosages due to the biphasic mode of action mentioned above) and require several weeks of chronic dosing before serotonin concentrations reach their maximal elevation (due to 1A autoreceptor desensitization) and full clinical benefits for conditions such as depression and anxiety are seen[72][73] (although other studies show an acute increase in 5-HT[74][75] which may account for initial worsening of symptoms in sensitive individuals[76]). For these reasons, selective serotonin releasing agents (SSRAs) such as MDAI and MMAI have been proposed as novel antidepressants with a putatively faster onset of action and improved effectiveness compared to current treatments.[72]
Similarly to SRAs, sufficiently high doses of 5-HT1A receptor agonists also bypass the 5-HT1A autoreceptor-mediated inhibition of serotonin release and therefore increase 5-HT1A postsynaptic receptor activation by directly agonizing the postsynaptic receptors in lieu of serotonin.
Ligands
The distribution of 5-HT1A receptors in the human brain may be imaged with the positron emission tomography using the radioligand [11C] WAY-100,635.[77] For example, one study has found increased 5-HT1A binding in type 2 diabetes.[78] Another PET study found a negative correlation between the amount of 5-HT1A binding in the raphe nuclei, hippocampus and neocortex and a self-reported tendency to have spiritual experiences.[79] Labeled with tritium, WAY-100,635 may also be used in autoradiography.[80]
Partial agonists
- 2C-B[81]
- 2C-E[81]
- 2C-T-2[81]
- 4C-T-2[81]
- 5-CT
- 5-MT
- 5-MeO-DiPT[81]
- 5-MeO-DMT
- 5-MeO-MiPT[81]
- 5-MeO-TMT[81]
- 6-F-DMT[81]
- Adatanserin
- αET
- Amphetamine
- αMT
- Aripiprazole
- Asenapine
- Bacoside
- Bay R 1531
- Befiradol
- Binospirone
- Brexpiprazole
- Bufotenin
- Buspirone (postsynaptic 5-HT1A)
- Cannabidiol[82]
- Cariprazine
- Clozapine
- cis-LSZ[81]
- Dihydroergotamine
- Dimethyltryptamine
- DiPT[81]
- DOET[81]
- DOI[81]
- DPT[81]
- Ebalzotan
- Eltoprazine
- EMDT[81]
- Ergotamine
- Etoperidone
- F-11,461
- F-12,826
- F-13,714
- F-14,679
- Flesinoxan
- Flibanserin
- Ginkgo biloba[83]
- Gepirone
- Haloperidol
- Lamotrigine
- Ipsapirone
- Limonene
- Lisuride
- Lurasidone
- LY-301,317
- Lysergic acid diethylamide (LSD)
- Mescaline[81]
- 3,4-Methylenedioxyamphetamine (MDA)[81]
- 3,4-Methylenedioxymethamphetamine (MDMA)[84]
- Methylphenidate
- Methysergide
- Naluzotan
- NBUMP
- Nefazodone
- Olanzapine
- Osemozotan (postsynaptic 5-HT1A)
- Perospirone
- Pyrimidinylpiperazine
- Piclozotan
- Psilocin
- Psilocybin
- Quetiapine
- Rauwolscine
- Roxindole
- RR-2B[81]
- RU-24,969
- S-15,535
- Sarizotan
- SS-2C[81]
- SSR-181,507
- Sunepitron
- Tandospirone
- Tiospirone
- Trazodone
- Trifluoromethylphenylpiperazine
- Trimethoxyamphetamine[81]
- Umespirone
- Urapidil
- Vilazodone
- Xaliproden
- Yohimbine[85][86][87][88]
- Zalospirone
- Ziprasidone
Full agonists
- 8-OH-DPAT[89]
- Alnespirone
- Buspirone (presynaptic 5-HT1A)
- Befiradol
- Tetrahydrocannabivarin (THCV)
- Eptapirone
- Lesopitron
- MKC-242
- LY-293,284
- Osemozotan (presynaptic 5-HT1A)
- Repinotan
- U-92,016-A
- Flibanserin
- Vortioxetine
Biased agonists
Antagonists
- Alprenolol
- AV-965
- Binospirone (postsynaptic 5-HT1A)
- BMY-7,378
- Cannabigerol
- Cyanopindolol
- Cyproheptadine
- Dotarizine
- Flopropione
- GR-46,611
- Iodocyanopindolol
- Isamoltane
- Lecozotan
- Mefway
- Methiothepin
- MPPF
- NAN-190
- Nebivolol
- Oxprenolol
- Pindobind
- Pindolol (presynaptic 5-HT1A)
- Propranolol
- Risperidone (weak)
- Robalzotan
- SB-649,915
- SDZ-216,525
- Spiperone
- Spiramide
- Spiroxatrine
- UH-301
- WAY-100,135
- WAY-100,635
- Xylamidine
Genetics
The 5-HT1A receptor is coded by the HTR1A gene. There are several human polymorphisms associated with this gene. A 2007 review listed 27 single nucleotide polymorphisms (SNP).[97] The most investigated SNPs are C-1019G (rs6295), C-1018G,[98] Ile28Val (rs1799921), Arg219Leu (rs1800044), and Gly22Ser (rs1799920).[97] Some of the other SNPs are Pro16Leu, Gly272Asp, and the synonymous polymorphism G294A (rs6294). These gene variants have been studied in relation to psychiatric disorders with no definitive results.[97]
Protein-protein interactions
The 5-HT1A receptor has been shown to interact with brain-derived neurotrophic factor (BDNF), which may play a major role in its regulation of mood and anxiety.[99][100]
See also
References
- GRCh38: Ensembl release 89: ENSG00000178394 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000021721 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Gilliam TC, Freimer NB, Kaufmann CA, Powchik PP, Bassett AS, Bengtsson U, Wasmuth JJ (November 1989). "Deletion mapping of DNA markers to a region of chromosome 5 that cosegregates with schizophrenia". Genomics. 5 (4): 940–4. doi:10.1016/0888-7543(89)90138-9. PMC 3154173. PMID 2591972.
- "Entrez Gene: HTR1A 5-hydroxytryptamine (serotonin) receptor 1A".
- Ito H, Halldin C, Farde L (January 1999). "Localization of 5-HT1A receptors in the living human brain using [carbonyl-11C]WAY-100635: PET with anatomic standardization technique". Journal of Nuclear Medicine. 40 (1): 102–9. PMID 9935065.
- Glennon RA, Dukat M, Westkaemper RB (2000-01-01). "Serotonin Receptor Subtypes and Ligands". American College of Neurophyscopharmacology. Archived from the original on 21 April 2008. Retrieved 2008-04-11.
- de Almeida J, Mengod G (October 2008). "Serotonin 1A receptors in human and monkey prefrontal cortex are mainly expressed in pyramidal neurons and in a GABAergic interneuron subpopulation: implications for schizophrenia and its treatment". Journal of Neurochemistry. 107 (2): 488–96. doi:10.1111/j.1471-4159.2008.05649.x. PMID 18761712. S2CID 23783438.
- Dabiré H (1991). "Central 5-hydroxytryptamine (5-HT) receptors in blood pressure regulation". Therapie. 46 (6): 421–9. PMID 1819150.
- Ramage AG (April 1991). "The mechanism of the sympathoinhibitory action of urapidil: role of 5-HT1A receptors". British Journal of Pharmacology. 102 (4): 998–1002. doi:10.1111/j.1476-5381.1991.tb12290.x. PMC 1917978. PMID 1855130.
- Kolassa N, Beller KD, Sanders KH (August 1989). "Involvement of brain 5-HT1A receptors in the hypotensive response to urapidil". The American Journal of Cardiology. 64 (7): 7D–10D. doi:10.1016/0002-9149(89)90688-7. PMID 2569265.
- Ootsuka Y, Blessing WW (February 2006). "Activation of 5-HT1A receptors in rostral medullary raphé inhibits cutaneous vasoconstriction elicited by cold exposure in rabbits". Brain Research. 1073–1074: 252–61. doi:10.1016/j.brainres.2005.12.031. PMID 16455061. S2CID 23178233.
- Rusyniak DE, Zaretskaia MV, Zaretsky DV, DiMicco JA (November 2007). "3,4-Methylenedioxymethamphetamine- and 8-hydroxy-2-di-n-propylamino-tetralin-induced hypothermia: role and location of 5-hydroxytryptamine 1A receptors". The Journal of Pharmacology and Experimental Therapeutics. 323 (2): 477–87. doi:10.1124/jpet.107.126169. PMID 17702902. S2CID 14197613.
- Yu Y, Ramage AG, Koss MC (April 2004). "Pharmacological studies of 8-OH-DPAT-induced pupillary dilation in anesthetized rats". European Journal of Pharmacology. 489 (3): 207–13. doi:10.1016/j.ejphar.2004.03.007. PMID 15087245.
- Prow MR, Martin KF, Heal DJ (December 1996). "8-OH-DPAT-induced mydriasis in mice: a pharmacological characterisation". European Journal of Pharmacology. 317 (1): 21–8. doi:10.1016/S0014-2999(96)00693-0. PMID 8982715.
- Fanciullacci M, Sicuteri R, Alessandri M, Geppetti P (March 1995). "Buspirone, but not sumatriptan, induces miosis in humans: relevance for a serotoninergic pupil control". Clinical Pharmacology and Therapeutics. 57 (3): 349–55. doi:10.1016/0009-9236(95)90161-2. PMID 7697953. S2CID 24512607.
- Cohn JB, Rickels K (1989). "A pooled, double-blind comparison of the effects of buspirone, diazepam and placebo in women with chronic anxiety". Current Medical Research and Opinion. 11 (5): 304–20. doi:10.1185/03007998909115213. PMID 2649317.
- Cryan JF, Redmond AM, Kelly JP, Leonard BE (May 1997). "The effects of the 5-HT1A agonist flesinoxan, in three paradigms for assessing antidepressant potential in the rat". European Neuropsychopharmacology. 7 (2): 109–14. doi:10.1016/S0924-977X(96)00391-4. PMID 9169298. S2CID 42048881.
- Parks CL, Robinson PS, Sibille E, Shenk T, Toth M (September 1998). "Increased anxiety of mice lacking the serotonin1A receptor". Proceedings of the National Academy of Sciences of the United States of America. 95 (18): 10734–9. Bibcode:1998PNAS...9510734P. doi:10.1073/pnas.95.18.10734. PMC 27964. PMID 9724773.
- Kennett GA, Dourish CT, Curzon G (February 1987). "Antidepressant-like action of 5-HT1A agonists and conventional antidepressants in an animal model of depression". European Journal of Pharmacology. 134 (3): 265–74. doi:10.1016/0014-2999(87)90357-8. PMID 2883013.
- Keller MB, Ruwe FJ, Janssens CJ, Sitsen JM, Jokinen R, Janczewski J (February 2005). "Relapse prevention with gepirone ER in outpatients with major depression". Journal of Clinical Psychopharmacology. 25 (1): 79–84. doi:10.1097/01.jcp.0000150221.53877.d9. PMID 15643103. S2CID 72677194.
- Invernizzi RW, Sacchetti G, Parini S, Acconcia S, Samanin R (August 2003). "Flibanserin, a potential antidepressant drug, lowers 5-HT and raises dopamine and noradrenaline in the rat prefrontal cortex dialysate: role of 5-HT(1A) receptors". British Journal of Pharmacology. 139 (7): 1281–8. doi:10.1038/sj.bjp.0705341. PMC 1573953. PMID 12890707.
- de Paulis T (January 2007). "Drug evaluation: PRX-00023, a selective 5-HT1A receptor agonist for depression". Current Opinion in Investigational Drugs. 8 (1): 78–86. PMID 17263189.
- Greenberg WM, Citrome L (May 2017). "Pharmacokinetics and Pharmacodynamics of Lurasidone Hydrochloride, a Second-Generation Antipsychotic: A Systematic Review of the Published Literature". Clinical Pharmacokinetics. 56 (5): 493–503. doi:10.1007/s40262-016-0465-5. PMID 27722855. S2CID 207485482.
- Stark AD, Jordan S, Allers KA, Bertekap RL, Chen R, Mistry Kannan T, et al. (February 2007). "Interaction of the novel antipsychotic aripiprazole with 5-HT1A and 5-HT 2A receptors: functional receptor-binding and in vivo electrophysiological studies". Psychopharmacology. 190 (3): 373–82. doi:10.1007/s00213-006-0621-y. PMID 17242925. S2CID 25349673.
- Wheeler Vega JA, Mortimer AM, Tyson PJ (May 2003). "Conventional antipsychotic prescription in unipolar depression, I: an audit and recommendations for practice". The Journal of Clinical Psychiatry. 64 (5): 568–74. doi:10.4088/JCP.v64n0512. PMID 12755661. Archived from the original on 20 June 2009.
- Donaldson ZR, Nautiyal KM, Ahmari SE, Hen R (June 2013). "Genetic approaches for understanding the role of serotonin receptors in mood and behavior". Current Opinion in Neurobiology. 23 (3): 399–406. doi:10.1016/j.conb.2013.01.011. PMC 3652904. PMID 23385115.
- Blier P, Abbott FV (January 2001). "Putative mechanisms of action of antidepressant drugs in affective and anxiety disorders and pain" (PDF). Journal of Psychiatry & Neuroscience. 26 (1): 37–43. PMC 1408043. PMID 11212592. Archived from the original (PDF) on 2016-03-06. Retrieved 2009-07-05.
- Morley KC, Arnold JC, McGregor IS (June 2005). "Serotonin (1A) receptor involvement in acute 3,4-methylenedioxymethamphetamine (MDMA) facilitation of social interaction in the rat". Progress in Neuro-Psychopharmacology & Biological Psychiatry. 29 (5): 648–57. doi:10.1016/j.pnpbp.2005.04.009. PMID 15908091. S2CID 24451268.
- Thompson MR, Callaghan PD, Hunt GE, Cornish JL, McGregor IS (May 2007). "A role for oxytocin and 5-HT(1A) receptors in the prosocial effects of 3,4 methylenedioxymethamphetamine ("ecstasy")". Neuroscience. 146 (2): 509–14. doi:10.1016/j.neuroscience.2007.02.032. PMID 17383105. S2CID 15617471.
- Gobbi G, Cassano T, Radja F, Morgese MG, Cuomo V, Santarelli L, et al. (April 2007). "Neurokinin 1 receptor antagonism requires norepinephrine to increase serotonin function". European Neuropsychopharmacology. 17 (5): 328–38. doi:10.1016/j.euroneuro.2006.07.004. PMID 16950604. S2CID 24350120.
- Baker KG, Halliday GM, Hornung JP, Geffen LB, Cotton RG, Törk I (1991). "Distribution, morphology and number of monoamine-synthesizing and substance P-containing neurons in the human dorsal raphe nucleus". Neuroscience. 42 (3): 757–75. doi:10.1016/0306-4522(91)90043-N. PMID 1720227. S2CID 23034680.
- Lucot JB (February 1994). "Antiemetic effects of flesinoxan in cats: comparisons with 8-hydroxy-2-(di-n-propylamino)tetralin". European Journal of Pharmacology. 253 (1–2): 53–60. doi:10.1016/0014-2999(94)90756-0. PMID 8013549.
- Oshima T, Kasuya Y, Okumura Y, Terazawa E, Dohi S (November 2002). "Prevention of nausea and vomiting with tandospirone in adults after tympanoplasty". Anesthesia and Analgesia. 95 (5): 1442–5, table of contents. doi:10.1097/00000539-200211000-00063. PMID 12401641. S2CID 26108853.
- Bardin L, Tarayre JP, Malfetes N, Koek W, Colpaert FC (April 2003). "Profound, non-opioid analgesia produced by the high-efficacy 5-HT(1A) agonist F 13640 in the formalin model of tonic nociceptive pain". Pharmacology. 67 (4): 182–94. doi:10.1159/000068404. PMID 12595749. S2CID 25882138.
- Colpaert FC (January 2006). "5-HT(1A) receptor activation: new molecular and neuroadaptive mechanisms of pain relief". Current Opinion in Investigational Drugs. 7 (1): 40–7. PMID 16425670.
- Blier P, Gobbi G, Haddjeri N, Santarelli L, Mathew G, Hen R (May 2004). "Impact of substance P receptor antagonism on the serotonin and norepinephrine systems: relevance to the antidepressant/anxiolytic response". Journal of Psychiatry & Neuroscience. 29 (3): 208–18. PMC 400690. PMID 15173897.
- Li Z, Ichikawa J, Dai J, Meltzer HY (June 2004). "Aripiprazole, a novel antipsychotic drug, preferentially increases dopamine release in the prefrontal cortex and hippocampus in rat brain". European Journal of Pharmacology. 493 (1–3): 75–83. doi:10.1016/j.ejphar.2004.04.028. PMID 15189766.
- Rollema H, Lu Y, Schmidt AW, Sprouse JS, Zorn SH (August 2000). "5-HT(1A) receptor activation contributes to ziprasidone-induced dopamine release in the rat prefrontal cortex". Biological Psychiatry. 48 (3): 229–37. doi:10.1016/S0006-3223(00)00850-7. PMID 10924666. S2CID 54398705.
- Rollema H, Lu Y, Schmidt AW, Zorn SH (November 1997). "Clozapine increases dopamine release in prefrontal cortex by 5-HT1A receptor activation". European Journal of Pharmacology. 338 (2): R3-5. doi:10.1016/S0014-2999(97)81951-6. PMID 9456005.
- Yoshino T, Nisijima K, Katoh S, Yui K, Nakamura M (April 2002). "Tandospirone potentiates the fluoxetine-induced increases in extracellular dopamine via 5-HT(1A) receptors in the rat medial frontal cortex". Neurochemistry International. 40 (4): 355–60. doi:10.1016/S0197-0186(01)00079-1. PMID 11792466. S2CID 41740125.
- Chojnacka-Wójcik E, Tatarczyńska E, Gołembiowska K, Przegaliński E (July 1991). "Involvement of 5-HT1A receptors in the antidepressant-like activity of gepirone in the forced swimming test in rats". Neuropharmacology. 30 (7): 711–7. doi:10.1016/0028-3908(91)90178-E. PMID 1681449. S2CID 46082801.
- Ogren SO, Eriksson TM, Elvander-Tottie E, D'Addario C, Ekström JC, Svenningsson P, et al. (December 2008). "The role of 5-HT(1A) receptors in learning and memory". Behavioural Brain Research. 195 (1): 54–77. doi:10.1016/j.bbr.2008.02.023. PMID 18394726. S2CID 140205386.
- Meltzer HY, Sumiyoshi T (December 2008). "Does stimulation of 5-HT(1A) receptors improve cognition in schizophrenia?". Behavioural Brain Research. 195 (1): 98–102. doi:10.1016/j.bbr.2008.05.016. PMID 18707769. S2CID 18455503.
- Harder JA, Ridley RM (February 2000). "The 5-HT1A antagonist, WAY 100 635, alleviates cognitive impairments induced by dizocilpine (MK-801) in monkeys". Neuropharmacology. 39 (4): 547–52. doi:10.1016/s0028-3908(99)00179-3. PMID 10728875. S2CID 43303149.
- Harder JA, Maclean CJ, Alder JT, Francis PT, Ridley RM (October 1996). "The 5-HT1A antagonist, WAY 100635, ameliorates the cognitive impairment induced by fornix transection in the marmoset". Psychopharmacology. 127 (3): 245–54. doi:10.1007/bf02246133. PMID 8912403. S2CID 10132368.
- Spreitzer H (August 13, 2008). "Neue Wirkstoffe - Lecozotan". Österreichische Apothekerzeitung (in German) (17/2007): 805.
- de Boer SF, Koolhaas JM (December 2005). "5-HT1A and 5-HT1B receptor agonists and aggression: a pharmacological challenge of the serotonin deficiency hypothesis". European Journal of Pharmacology. 526 (1–3): 125–39. doi:10.1016/j.ejphar.2005.09.065. PMID 16310183.
- Olivier B, Mos J, Rasmussen D (1990). "Behavioural pharmacology of the serenic, eltoprazine". Drug Metabolism and Drug Interactions. 8 (1–2): 31–83. doi:10.1515/DMDI.1990.8.1-2.31. PMID 2091890. S2CID 27279453.
- Winstanley CA, Theobald DE, Dalley JW, Robbins TW (April 2005). "Interactions between serotonin and dopamine in the control of impulsive choice in rats: therapeutic implications for impulse control disorders". Neuropsychopharmacology. 30 (4): 669–82. doi:10.1038/sj.npp.1300610. PMID 15688093.
- Tomkins DM, Higgins GA, Sellers EM (June 1994). "Low doses of the 5-HT1A agonist 8-hydroxy-2-(di-n-propylamino)-tetralin (8-OH DPAT) increase ethanol intake". Psychopharmacology. 115 (1–2): 173–9. doi:10.1007/BF02244769. PMID 7862892. S2CID 38012716.
- Müller CP, Carey RJ, Huston JP, De Souza Silva MA (February 2007). "Serotonin and psychostimulant addiction: focus on 5-HT1A-receptors". Progress in Neurobiology. 81 (3): 133–78. doi:10.1016/j.pneurobio.2007.01.001. PMID 17316955. S2CID 42788995.
- Carey RJ, DePalma G, Damianopoulos E, Shanahan A, Müller CP, Huston JP (February 2005). "Evidence that the 5-HT1A autoreceptor is an important pharmacological target for the modulation of cocaine behavioral stimulant effects". Brain Research. 1034 (1–2): 162–71. doi:10.1016/j.brainres.2004.12.012. PMID 15713268. S2CID 28356741.
- Fernández-Guasti A, Rodríguez-Manzo G (January 1997). "8-OH-DPAT and male rat sexual behavior: partial blockade by noradrenergic lesion and sexual exhaustion". Pharmacology, Biochemistry, and Behavior. 56 (1): 111–6. doi:10.1016/S0091-3057(96)00165-7. PMID 8981617. S2CID 26063338.
- Haensel SM, Slob AK (July 1997). "Flesinoxan: a prosexual drug for male rats". European Journal of Pharmacology. 330 (1): 1–9. doi:10.1016/S0014-2999(97)00170-2. PMID 9228408.
- Simon P, Guardiola B, Bizot-Espiard J, Schiavi P, Costentin J (1992). "5-HT1A receptor agonists prevent in rats the yawning and penile erections induced by direct dopamine agonists". Psychopharmacology. 108 (1–2): 47–50. doi:10.1007/BF02245284. PMID 1357709. S2CID 22385029.
- Millan MJ, Perrin-Monneyron S (March 1997). "Potentiation of fluoxetine-induced penile erections by combined blockade of 5-HT1A and 5-HT1B receptors". European Journal of Pharmacology. 321 (3): R11-3. doi:10.1016/S0014-2999(97)00050-2. PMID 9085055.
- Ebenezer IS, Arkle MJ, Tite RM (May 2007). "8-Hydroxy-2-(di-n-propylamino)-tetralin inhibits food intake in fasted rats by an action at 5-HT1A receptors". Methods and Findings in Experimental and Clinical Pharmacology. 29 (4): 269–72. doi:10.1358/mf.2007.29.4.1075362. PMID 17609739.
- Monti JM, Jantos H (September 1992). "Dose-dependent effects of the 5-HT1A receptor agonist 8-OH-DPAT on sleep and wakefulness in the rat". Journal of Sleep Research. 1 (3): 169–175. doi:10.1111/j.1365-2869.1992.tb00033.x. PMID 10607047. S2CID 27917774.
- Ansseau M, Pitchot W, Gonzalez Moreno A, Wauthy J, Papart P (2004). "Pilot study of flesinoxan, a 5-HT1A agonist, in major depression: Effects on sleep REM latency and body temperature". Human Psychopharmacology: Clinical and Experimental. 8 (4): 279–283. doi:10.1002/hup.470080407. S2CID 145758823. Archived from the original on 2012-12-17.
- Meyer LC, Fuller A, Mitchell D (February 2006). "Zacopride and 8-OH-DPAT reverse opioid-induced respiratory depression and hypoxia but not catatonic immobilization in goats". American Journal of Physiology. Regulatory, Integrative and Comparative Physiology. 290 (2): R405-13. doi:10.1152/ajpregu.00440.2005. PMID 16166206. S2CID 224414.
- Van de Kar LD, Levy AD, Li Q, Brownfield MS (July 1998). "A comparison of the oxytocin and vasopressin responses to the 5-HT1A agonist and potential anxiolytic drug alnespirone (S-20499)". Pharmacology, Biochemistry, and Behavior. 60 (3): 677–83. doi:10.1016/S0091-3057(98)00025-2. PMID 9678651. S2CID 27817530.
- Lorens SA, Van de Kar LD (April 1987). "Differential effects of serotonin (5-HT1A and 5-HT2) agonists and antagonists on renin and corticosterone secretion". Neuroendocrinology. 45 (4): 305–10. doi:10.1159/000124754. PMID 2952898.
- Koenig JI, Gudelsky GA, Meltzer HY (May 1987). "Stimulation of corticosterone and beta-endorphin secretion in the rat by selective 5-HT receptor subtype activation". European Journal of Pharmacology. 137 (1): 1–8. doi:10.1016/0014-2999(87)90175-0. PMID 2956114.
- Pitchot W, Wauthy J, Legros JJ, Ansseau M (March 2004). "Hormonal and temperature responses to flesinoxan in normal volunteers: an antagonist study". European Neuropsychopharmacology. 14 (2): 151–5. doi:10.1016/S0924-977X(03)00108-1. PMID 15013031. S2CID 19082134.
- Navinés R, Martín-Santos R, Gómez-Gil E, Martínez de Osaba MJ, Gastó C (December 2008). "Interaction between serotonin 5-HT1A receptors and beta-endorphins modulates antidepressant response". Progress in Neuro-Psychopharmacology & Biological Psychiatry. 32 (8): 1804–9. doi:10.1016/j.pnpbp.2008.07.021. PMID 18725263. S2CID 37943722.
- Hjorth S, Bengtsson HJ, Kullberg A, Carlzon D, Peilot H, Auerbach SB (June 2000). "Serotonin autoreceptor function and antidepressant drug action". Journal of Psychopharmacology. 14 (2): 177–85. doi:10.1177/026988110001400208. PMID 10890313. S2CID 33440228.
- Briley M, Moret C (October 1993). "Neurobiological mechanisms involved in antidepressant therapies". Clinical Neuropharmacology. 16 (5): 387–400. doi:10.1097/00002826-199310000-00002. PMID 8221701.
- Starr KR, Price GW, Watson JM, Atkinson PJ, Arban R, Melotto S, et al. (October 2007). "SB-649915-B, a novel 5-HT1A/B autoreceptor antagonist and serotonin reuptake inhibitor, is anxiolytic and displays fast onset activity in the rat high light social interaction test". Neuropsychopharmacology. 32 (10): 2163–72. doi:10.1038/sj.npp.1301341. PMID 17356576.
- Rothman RB, Baumann MH (2006). "Therapeutic potential of monoamine transporter substrates". Current Topics in Medicinal Chemistry. 6 (17): 1845–59. doi:10.2174/156802606778249766. PMID 17017961. Archived from the original on 2017-03-26. Retrieved 2019-04-30.
- Scorza C, Silveira R, Nichols DE, Reyes-Parada M (July 1999). "Effects of 5-HT-releasing agents on the extracellullar hippocampal 5-HT of rats. Implications for the development of novel antidepressants with a short onset of action". Neuropharmacology. 38 (7): 1055–61. doi:10.1016/S0028-3908(99)00023-4. PMID 10428424. S2CID 13714807.
- Marona-Lewicka D, Nichols DE (July 1998). "Drug discrimination studies of the interoceptive cues produced by selective serotonin uptake inhibitors and selective serotonin releasing agents". Psychopharmacology. 138 (1): 67–75. doi:10.1007/s002130050646. PMID 9694528. S2CID 32698247. Archived from the original on 2002-01-12. Retrieved 2009-07-05.
- Fuller RW (1994). "Uptake inhibitors increase extracellular serotonin concentration measured by brain microdialysis". Life Sciences. 55 (3): 163–7. doi:10.1016/0024-3205(94)00876-0. PMID 8007758.
- Rutter JJ, Auerbach SB (June 1993). "Acute uptake inhibition increases extracellular serotonin in the rat forebrain". The Journal of Pharmacology and Experimental Therapeutics. 265 (3): 1319–24. PMID 7685386.
- Bigos KL, Pollock BG, Aizenstein HJ, Fisher PM, Bies RR, Hariri AR (December 2008). "Acute 5-HT reuptake blockade potentiates human amygdala reactivity". Neuropsychopharmacology. 33 (13): 3221–5. doi:10.1038/npp.2008.52. PMC 2858321. PMID 18463627.
- Pike VW, McCarron JA, Lammerstma AA, Hume SP, Poole K, Grasby PM, et al. (September 1995). "First delineation of 5-HT1A receptors in human brain with PET and [11C]WAY-100635". European Journal of Pharmacology. 283 (1–3): R1-3. doi:10.1016/0014-2999(95)00438-Q. PMID 7498295.
- Price JC, Kelley DE, Ryan CM, Meltzer CC, Drevets WC, Mathis CA, et al. (February 2002). "Evidence of increased serotonin-1A receptor binding in type 2 diabetes: a positron emission tomography study". Brain Research. 927 (1): 97–103. doi:10.1016/S0006-8993(01)03297-8. PMID 11814436. S2CID 32547663.
- Borg J, Andrée B, Soderstrom H, Farde L (November 2003). "The serotonin system and spiritual experiences". The American Journal of Psychiatry. 160 (11): 1965–9. doi:10.1176/appi.ajp.160.11.1965. PMID 14594742. S2CID 5911066.
- Burnet PW, Eastwood SL, Harrison PJ (June 1997). "[3H]WAY-100635 for 5-HT1A receptor autoradiography in human brain: a comparison with [3H]8-OH-DPAT and demonstration of increased binding in the frontal cortex in schizophrenia". Neurochemistry International. 30 (6): 565–74. doi:10.1016/S0197-0186(96)00124-6. PMID 9152998. S2CID 21135585.
- Ray TS (February 2010). "Psychedelics and the human receptorome". PLOS ONE. 5 (2): e9019. Bibcode:2010PLoSO...5.9019R. doi:10.1371/journal.pone.0009019. PMC 2814854. PMID 20126400.
- Russo EB, Burnett A, Hall B, Parker KK (August 2005). "Agonistic properties of cannabidiol at 5-HT1a receptors". Neurochemical Research. 30 (8): 1037–43. doi:10.1007/s11064-005-6978-1. PMID 16258853. S2CID 207222631.
- Winter JC, Timineri D (March 1999). "The discriminative stimulus properties of EGb 761, an extract of Ginkgo biloba". Pharmacology, Biochemistry, and Behavior. 62 (3): 543–7. doi:10.1016/S0091-3057(98)00190-7. PMID 10080249. S2CID 23055772.
- Oeri, HE (2020). "Beyond ecstasy: alternative entactogens to 3,4-methylenedioxymethamphetamine with potential applications in psychotherapy". Journal of Psychopharmacology. 35 (5): 512–536. doi:10.1177/0269881120920420. PMC 8155739. PMID 32909493.
- Arthur JM, Casañas SJ, Raymond JR (June 1993). "Partial agonist properties of rauwolscine and yohimbine for the inhibition of adenylyl cyclase by recombinant human 5-HT1A receptors". Biochemical Pharmacology. 45 (11): 2337–41. doi:10.1016/0006-2952(93)90208-E. PMID 8517875.
- Kaumann AJ (June 1983). "Yohimbine and rauwolscine inhibit 5-hydroxytryptamine-induced contraction of large coronary arteries of calf through blockade of 5 HT2 receptors". Naunyn-Schmiedeberg's Archives of Pharmacology. 323 (2): 149–54. doi:10.1007/BF00634263. PMID 6136920. S2CID 23251900.
- Baxter GS, Murphy OE, Blackburn TP (May 1994). "Further characterization of 5-hydroxytryptamine receptors (putative 5-HT2B) in rat stomach fundus longitudinal muscle". British Journal of Pharmacology. 112 (1): 323–31. doi:10.1111/j.1476-5381.1994.tb13072.x. PMC 1910288. PMID 8032658.
- "Yohimbine (PIM 567)". Inchem.org. Retrieved 2013-05-26.
- Winsauer PJ, Rodriguez FH, Cha AE, Moerschbaecher JM (January 1999). "Full and partial 5-HT1A receptor agonists disrupt learning and performance in rats" (PDF). The Journal of Pharmacology and Experimental Therapeutics. 288 (1): 335–47. PMID 9862788.
- Pytka K, Głuch-Lutwin M, Żmudzka E, Sałaciak K, Siwek A, Niemczyk K, et al. (2018). "1A Receptor Ligand With Anxiolytic-Like Activity, Preferentially Activates ß-Arrestin Signaling". Frontiers in Pharmacology. 9: 1146. doi:10.3389/fphar.2018.01146. PMC 6209770. PMID 30410441.
- Sniecikowska J, Gluch-Lutwin M, Bucki A, Więckowska A, Siwek A, Jastrzebska-Wiesek M, et al. (March 2019). "1A Receptor-Biased Agonists with Robust Antidepressant-like Activity". Journal of Medicinal Chemistry. 62 (5): 2750–2771. doi:10.1021/acs.jmedchem.9b00062. PMID 30721053.
- Ignarro LJ (June 2008). "Different pharmacological properties of two enantiomers in a unique beta-blocker, nebivolol". Cardiovascular Therapeutics. 26 (2): 115–34. doi:10.1111/j.1527-3466.2008.00044.x. PMID 18485134.
- Prasad R, Paila YD, Chattopadhyay A (December 2009). "Membrane cholesterol depletion enhances ligand binding function of human serotonin1A receptors in neuronal cells". Biochem Biophys Res Commun. 390 (1): 93–6. doi:10.1016/j.bbrc.2009.09.072. PMID 19781522.
- Thomas EA, Carson MJ, Neal MJ, Sutcliffe JG (1997). "Unique allosteric regulation of 5-hydroxytryptamine receptor-mediated signal transduction by oleamide". Proc Natl Acad Sci U S A. 94 (25): 14115–9. doi:10.1073/pnas.94.25.14115. PMC 28442. PMID 9391162.
- Satała G, Duszyńska B, Lenda T, Nowak G, Bojarski AJ (2018). "Allosteric Inhibition of Serotonin 5-HT7 Receptors by Zinc Ions". Mol Neurobiol. 55 (4): 2897–2910. doi:10.1007/s12035-017-0536-0. PMC 5842505. PMID 28455702.
- Yano H, Adhikari P, Naing S, Hoffman AF, Baumann MH, Lupica CR, Shi L (2020). "Positive Allosteric Modulation of the 5-HT1A Receptor by Indole-Based Synthetic Cannabinoids Abused by Humans". ACS Chem Neurosci. 11 (10): 1400–1405. doi:10.1021/acschemneuro.0c00034. PMC 8275447. PMID 32324370.
- Drago A, Ronchi DD, Serretti A (August 2008). "5-HT1A gene variants and psychiatric disorders: a review of current literature and selection of SNPs for future studies". The International Journal of Neuropsychopharmacology. 11 (5): 701–21. doi:10.1017/S1461145707008218. PMID 18047755.
- Wu S, Comings DE (June 1999). "A common C-1018G polymorphism in the human 5-HT1A receptor gene". Psychiatric Genetics. 9 (2): 105–6. doi:10.1097/00041444-199906000-00010. PMID 10412191.
- Anttila S, Huuhka K, Huuhka M, Rontu R, Hurme M, Leinonen E, Lehtimäki T (2007). "Interaction between 5-HT1A and BDNF genotypes increases the risk of treatment-resistant depression". Journal of Neural Transmission. 114 (8): 1065–8. doi:10.1007/s00702-007-0705-9. PMID 17401528. S2CID 19373406.
- Guiard BP, David DJ, Deltheil T, Chenu F, Le Maître E, Renoir T, et al. (February 2008). "Brain-derived neurotrophic factor-deficient mice exhibit a hippocampal hyperserotonergic phenotype". The International Journal of Neuropsychopharmacology. 11 (1): 79–92. doi:10.1017/S1461145707007857. PMID 17559709.
- Renner U, Zeug A, Woehler A, Niebert M, Dityatev A, Dityateva G, et al. (May 2012). "Heterodimerization of serotonin receptors 5-HT1A and 5-HT7 differentially regulates receptor signalling and trafficking" (PDF). Journal of Cell Science. 125 (Pt 10): 2486–99. doi:10.1242/jcs.101337. hdl:11858/00-001M-0000-000F-A828-A. PMID 22357950. S2CID 970339.
- Salim K, Fenton T, Bacha J, Urien-Rodriguez H, Bonnert T, Skynner HA, et al. (May 2002). "Oligomerization of G-protein-coupled receptors shown by selective co-immunoprecipitation". The Journal of Biological Chemistry. 277 (18): 15482–5. doi:10.1074/jbc.M201539200. PMID 11854302.
External links
- "5-HT1A". IUPHAR Database of Receptors and Ion Channels. International Union of Basic and Clinical Pharmacology.
- Human HTR1A genome location and HTR1A gene details page in the UCSC Genome Browser.
This article incorporates text from the United States National Library of Medicine, which is in the public domain.
| Main articles and pharmaceuticals |
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Neuropharmacology |
| ||||||||
| Active metabolites | |||||||||
| Related articles |
| ||||||||
| |||||||||